Chromalveolata
| Chromalveolata | |||
|---|---|---|---|
 | |||
| Kloksgewijs van links naar rechts: een haptofyt; diatomeeën; een waterschimmel; een cryptofyt; en Macrocystis, een bruinwier | |||
| Taxonomische indeling | |||
| |||
| Supergroep | |||
| Chromalveolata Cavalier-Smith, 1999[1] (niet monofyletisch) | |||
| Afbeeldingen op | |||
| Chromalveolata op | |||
| |||
De Chromalveolata vormen een omvangrijke en diverse groep van eukaryoten, die lange tijd werd beschouwd als een aparte supergroep binnen de biologische taxonomie. De Chromalveolata werd voorgesteld in 1999 door Thomas Cavalier-Smith,[1] een verfijning op het door hem geïntroduceerde rijk Chromista. Het idee was dat de Chromista en Alveolata een gemeenschappelijke fotosynthetische voorouder zouden delen.[2]
Het taxon Chromalveolata bevatte verschillende groepen fotosynthetiserende algen, zoals bruinwieren, dinoflagellaten, diatomeeën en Apicomplexa. Een definiërend aspect van de Chromalveolata was de oorsprong via secundaire endosymbiose: alle chromalveolaten zouden afstammen van een voorouderlijke eukaryote cel met twee flagellen (bikont) die een eencellige roodwier had opgenomen als endosymbiont.[3] Een gemeenschappelijk kenmerk was de aanwezigheid van chlorofyl c.
Later onderzoek naar moleculaire fylogenie van eukaryoten maakte duidelijk dat de Chromalveolata geen evolutionaire groep is, maar polyfyletisch.[4] De plastiden zijn in de loop van de evolutie meerdere keren onafhankelijk van elkaar verworven, en dus niet afkomstig uit een enkele voorouder.[5][6] Hierdoor werd het taxon in latere indelingen verlaten, en vonden de voormalige chromalveolaten een plaats in nieuw gedefinieerde supergroepen, zoals de SAR-clade en Haptista.
Taxonomie
Groepen en classificatie
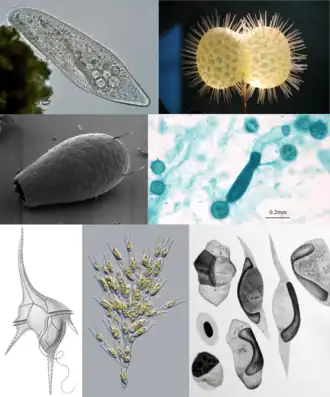
Het merendeel van de organismen die tot de chromalveolaten werden gerekend, werden in de twintigste eeuw ondergebracht in het rijk Protista. De waterschimmels (Oomycota) werden meestal in het rijk van de schimmels ingedeeld en de bruinwieren in het plantenrijk. Op basis van celbiologische kenmerken werden deze uiteenlopende organismen door de bioloog Thomas Cavalier-Smith samengevoegd onder de naam Chromista. In 1999 werd door hem voorgesteld dat Chromista samen met Alveolata één evolutionaire groep vormen: de Chromalveolata.[1]
In 2005 werd Chromalveolata als 'supergroep' opgenomen in een uitgebreide classificatie van eukaryoten die destijds een taxonomische consensus weerspiegelde.[7] De Chromalveolata werd beschouwd als een van de zes grote clades van eukaryoten. Hoewel de groep niet overal als formeel taxon werd erkend, werd het soms als een apart rijk behandeld. De Chromalveolata zou vier hoofdlijnen omvatten: Cryptophyta, Haptophyta, Stramenopiles en Alveolata.
Later in de jaren 2000 begon twijfel te ontstaan over de veronderstelling dat Chromalveolata een monofyletische groep vormt. Al in 2005 werd gewezen op het ontbreken van sterke argumenten voor een gemeenschappelijke voorouder.[8] Een overzichtsartikel uit 2006 stelde vergelijkbare problemen vast bij meerdere van de toen onderscheiden zes eukaryote supergroepen, waaronder de Chromalveolata.[9] Vanaf 2012 ontstond consensus onder biologen dat de groep niet monofyletisch is.[10] De vier oorspronkelijke subgroepen bleken tot ten minste twee verschillende evolutionaire lijnen te behoren: Stramenopiles en Alveolata vormen samen met de Rhizaria de SAR-supergroep. De Cryptophyta en Haptophyta werden elders geplaatst.
Huidige fylogenie
De evolutionaire relaties van fotosynthetische eukaryoten en verwante protisten is nog niet geheel opgehelderd. Dankzij uitgebreide bemonstering van vele soorten protisten en verfijning in fylogenetische analyses is in de loop der tijd een steeds sterker raamwerk ontstaan van de hogere clades.[11] De twee grote subclades, Archaeplastida en SAR, zijn relatief goed gekarakteriseerd en robuust monofyletisch.[12] Haptista en Cryptista, twee groepen waarvan aanvankelijk werd gedacht dat ze verwanten van elkaar waren, bleken later verder van elkaar af te staan.[13] Onderstaande fylogenie is gebaseerd op genoomanalyses van de jaren 2020.[14][11][15]
| Diaphoretickes |
| |||||||||||||||||||||||||||||||||
Eigenschappen
De meeste Chromalveolata hebben een autotrofe levenswijze en voorzien zichzelf van energie via fotosynthese. Alle fotosynthetiserende chromalveolaten gebruiken chlorofyl a en c en vele gebruiken daarnaast andere pigmenten. Ook diverse niet-fotosynthetische soorten behoren tot de traditionele Chromalveolata, zoals de Ciliophora en waterschimmels.
Morfologie

Er zijn niet veel morfologische eigenschappen die alle chromalveolaten delen. Elke ondergroep heeft zijn eigen unieke eigenschappen, zoals de alveoli van de Alveolata, de haptonema van de Haptophyta, het ejectosoom van de Cryptophyta en de twee verschillende flagella bij de Heterokontophyta. Geen van al deze eigenschappen is bij alle groepen aanwezig.
De meeste chromalveolaten hebben een celwand bestaande uit cellulose. De gezamenlijke oorsprong van de plastide (onder andere gebaseerd op membraantopologie) werd door Thomas Cavalier-Smith samen met andere celbiologische aspecten gezien als een belangrijk verenigend kenmerk.[16] Aan de hand van moleculair onderzoek is het echter duidelijk geworden dat plastiden meerdere keren onafhankelijk zijn verkregen of doorgegeven in de loop van de evolutie.[4][17]
Ecologische rol
Chromalveolaten spelen soms een belangrijke rol in ecosystemen. Sommige kunnen zeer schadelijk zijn. Dinoflagellaten kunnen algenbloei veroorzaken die vis- en oesterpopulaties in zee kunnen uitroeien. Apicomplexa omvatten een aantal belangrijke parasieten voor mens en dier. Waterschimmels zorgen voor ziekten bij planten; zo veroorzaakte de waterschimmel Phytophthora infestans de Grote Aardappelhongersnood in Ierland tussen 1845 en 1850.
Diatomeeën zijn een van de grootste bronnen van fotosynthese en produceren een groot deel van de zuurstof in de atmosfeer. Mariene bruinwieren, vooral kelp, creëren onder water wouden waarin veel soorten een habitat vinden.
Zie ook
Referenties
- 1 2 3 (en) Cavalier‐smith T. (1999). Principles of Protein and Lipid Targeting in Secondary Symbiogenesis: Euglenoid, Dinoflagellate, and Sporozoan Plastid Origins and the Eukaryote Family Tree. Journal of Eukaryotic Microbiology 46 (4): 347-366. DOI: 10.1111/j.1550-7408.1999.tb04614.x.
- ↑ (en) Green BR. (2011). After the primary endosymbiosis: an update on the chromalveolate hypothesis and the origins of algae with Chl c. Photosynthesis Research 107 (1): 103-115. DOI: 10.1007/s11120-010-9584-2.
- ↑ (en) Keeling PJ. (2009). Chromalveolates and the Evolution of Plastids by Secondary Endosymbiosis. Journal of Eukaryotic Microbiology 56 (1): 1-8. DOI: 10.1111/j.1550-7408.2008.00371.x.
- 1 2 3 (en) Burki F, Roger AJ, Brown MW, Simpson AG. (2020). The New Tree of Eukaryotes. Trends in Ecology & Evolution 35 (1): 43-55. DOI: 10.1016/j.tree.2019.08.008.
- ↑ (en) Burki F, Kaplan M, Tikhonenkov DV, Zlatogursky V, Minh BQ, Radaykina LV, Smirnov A. (2016). Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista. Proceedings of the Royal Society B: Biological Sciences 283 (1823): 20152802. DOI: 10.1098/rspb.2015.2802.
- ↑ (en) Pietluch F, Mackiewicz P, Ludwig K, Gagat P. (2024). A New Model and Dating for the Evolution of Complex Plastids of Red Alga Origin. Genome Biology and Evolution 16 (9). DOI: 10.1093/gbe/evae192.
- ↑ (en) Adl SM, Simpson AGB, Farmer MA, Andersen RA, Anderson OR, Barta JR, Bowser SS. (2005). The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. The Journal of Eukaryotic Microbiology 52 (5): 399-451. DOI: 10.1111/j.1550-7408.2005.00053.x.
- ↑ (en) Harper JT, Waanders E, Keeling PJ. (2005). On the monophyly of chromalveolates using a six-protein phylogeny of eukaryotes. International Journal of Systematic and Evolutionary Microbiology 55 (1): 487-496. DOI: 10.1099/ijs.0.63216-0.
- ↑ (en) Parfrey LW, Barbero E, Lasser E, Dunthorn M, Bhattacharya D, Patterson DJ, Katz LA. (2006). Evaluating Support for the Current Classification of Eukaryotic Diversity. PLoS Genetics 2 (12): e220. DOI: 10.1371/journal.pgen.0020220.
- ↑ (en) Adl SM, Simpson AGB, Lane CE, Lukeš J, Bass D, Bowser SS, Brown MW. (2012). The Revised Classification of Eukaryotes. Journal of Eukaryotic Microbiology 59 (5): 429-514. DOI: 10.1111/j.1550-7408.2012.00644.x.
- 1 2 (en) Torruella G, Galindo L, Moreira D, López-García P. (2025). Phylogenomics of neglected flagellated protists supports a revised eukaryotic tree of life. Current Biology 35 (1): 198–207.e4. PMID 39642877. DOI: 10.1016/j.cub.2024.10.075.
- ↑ (en) Adl SM, Bass D, Lane CE, LukeŠ J, Schoch CL, Smirnov A, Agatha S. (2019). Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. Journal of Eukaryotic Microbiology 66 (1): 4-119. DOI: 10.1111/jeu.12691.
- ↑ (en) Fabien B, Noriko O, Pombert J, Keeling P. (2012). The evolutionary history of haptophytes and cryptophytes: phylogenomic evidence for separate origins. Proceedings of the Royal Society of London B: Biological Sciences 279 (1736): 2246–2254. PMID 22298847. DOI: 10.1098/rspb.2011.2301.
- ↑ (en) Schön ME, Zlatogursky V, Singh R. (2021). Single cell genomics reveals plastid-lacking Picozoa are close relatives of red algae. Nature Communications 12 (1). PMID 34789758. DOI: 10.1038/s41467-021-26918-0.
- ↑ (en) Valt M, Panek T, Mirzoyan S, Tice AK, Jones RE, Dohnalek V. (2025). Rare microbial relict sheds light on an ancient eukaryotic supergroup. Nature. DOI: 10.1038/s41586-025-09750-0.
- ↑ (en) Cavalier-Smith T. (2010). Kingdoms Protozoa and Chromista and the eozoan root of the eukaryotic tree. Biology Letters 6 (3): 342-345. DOI: 10.1098/rsbl.2009.0948.
- ↑ (en) Strassert JFH, Irisarri I, Williams TA, Burki F. (2021). A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids. Nature Communications 12 (1). DOI: 10.1038/s41467-021-22044-z.
Externe link
- International Society of Protistologists, officiële website