Folliculaire dendritische cel
-associated_meningeal_lymphatic_system_and_the_human_immunodeficiency_virus_(HIV)_reservoir_in_the_cervical_lymph_nodes_(CLNs)._Cropped_B.jpg)
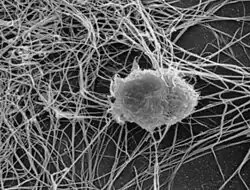
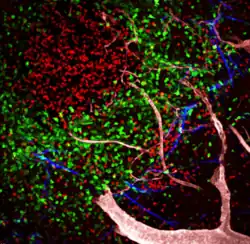
Folliculaire dendritische cellen zijn witte bloedcellen van het aangeboren immuunsysteem van veel zoogdieren die worden aangetroffen in primaire en secundaire lymfeklieren van de B-celgebieden van het lymfoïde weefsel.[1][2][3] De cortex van de lymfeklier bestaat uit groepen van voornamelijk geïnactiveerde B-cellen, follikels genaamd. Bij mensen hebben folliculaire dendritische cellen membraanachtige uitsteeksels.
In tegenstelling tot dendritische cellen zijn folliculaire dendritische cellen niet afkomstig van de hematopoëtische stamcel uit het beenmerg, maar zijn ze van mesenchymale oorsprong.[4] Mogelijke functies van folliculaire dendritische cellen zijn onder meer: het organiseren van de cellen en microarchitectuur van lymfoïde weefsel, het vangen van antigeen ter ondersteuning van B-cellen, het bevorderen van het verwijderen van afval uit kiemcentra en het beschermen tegen auto-immuniteit. Kiemcentra zijn tijdelijk gevormde structuren binnen de follikels.
Ziekteprocessen waaraan folliculaire dendritische cellen kunnen bijdragen, zijn onder andere primaire folliculaire dendritische celtumoren, chronische ontstekingsaandoeningen, de ontwikkeling van HIV-1-infecties en neuro-invasieve scrapie.
Locatie en moleculaire markers
Folliculaire dendritische cellen zijn een niet-migrerende populatie die wordt aangetroffen in primaire en secundaire follikels van de B-celgebieden van lymfeklieren, milt en mucosa-geassocieerd lymfoïde weefsel (slijmvlieslymfocyten). Ze vormen een stabiel netwerk dankzij intercellulaire verbindingen tussen folliculaire dendritische cellen-uitlopers en nauwe interactie met folliculaire B-cellen.[5][6] Het netwerk van folliculaire dendritische cellen vormt doorgaans het centrum van de follikel en strekt zich niet uit van het follikel naar de interfolliculaire regio's of de T-celzone. Verondersteld wordt dat deze scheiding van de plaatsen waar de eerste antigeenverwerking en het vangen plaatsvindt, een beschermde omgeving biedt waarin geopsoniseerde antigenen langdurig kunnen worden weergegeven zonder te worden geproteolyseerd of verwijderd door fagocyterende cellen. Opsoninen zijn extracellulaire eiwitten die, wanneer ze aan stoffen of cellen gebonden zijn, fagocyten aanzetten tot fagocytose van de stoffen of cellen waaraan de opsoninen gebonden zijn.
Folliculaire dendritische cellen hebben een hoge expressie van de complementreceptoren CR1 en CR2 (respectievelijk CD35 en CD21) en de Fc-receptor FcγRIIb (CD32). Verdere folliculaire dendritische cellen-specifieke moleculaire merkers zijn FDC-M1, FDC-M2 en C4.[7] In tegenstelling tot andere dendritische cellen en macrofagen, missen folliculaire dendritische cellen MHC klasse II antigeenmoleculen en brengen ze weinig patroonherkenningsreceptoren tot expressie, waardoor ze weinig vermogen hebben om niet-geopsoniseerde antigenen te vangen.[5]
Ontwikkeling
Folliculaire dendritische cellen ontwikkelen zich uit vermoedelijke mesenchymale voorlopercellen.[7] Modellen met muizen met ernstige gecombineerde immunodeficiëntie tonen aan dat deze voorlopercellen kunnen worden overgedragen op ontvangers met beenmergallotransplantaties. In dat geval kunnen de folliculaire dendritische cel-netwerken van zowel donoren als ontvangers later worden aangetroffen in de lymfoïde compartimenten van de ontvanger.[8] Interactie tussen folliculaire dendritische voorlopercellen en lymfoïde cellen, gemedieerd door TNF-α en TNF-β, is cruciaal voor de normale ontwikkeling en het behoud van folliculaire dendritische cellen. TNF-α bindt aan de TNFRI-receptor, terwijl tumornecrosefactor βinterageert met de TNF-β-receptor die tot expressie komt bij folliculaire dendritische voorlopercellen. Bij muizen zonder B-cellen, of met een geblokkeerde productie van TNF-α en TNF-β, ontbreken cellen met een folliculaire dendritische cel-fenotype.[9][10]
Functies
Organiserende lymfoïde microarchitectuur
In normaal lymfoïde weefsel migreren recirculerende rustende B-cellen door de folliculaire dendritische cel-netwerken, terwijl antigeen-geactiveerde B-cellen worden onderschept en klonale expansie ondergaan binnen de folliculaire dendritische cel-netwerken, waardoor kiemcentra ontstaan. Kiemcentra behoren tot de belangrijkste producenten van de chemokine CXCL13, die lymfoïde cellen aantrekt en organiseert.[11]
Antigeenvangende, geheugen-B-celondersteuning
Folliculaire dendritische cel-receptoren CR1, CR2 en FcγRIIb vangen antigeen op dat is geopsoniseerd door complement of antilichamen. Deze antigenen worden vervolgens opgenomen in een niet-afbrekend cyclisch endosomaalcompartiment voor latere presentatie aan B-cellen.[12] Om geselecteerd te worden als toekomstige geheugencel, moeten GK-B-cellen binden aan het antigeen dat door de folliculaire dendritische cellen wordt gevormd, anders gaan ze in apoptose.
Verwijdering van afbraakproducten
Door de uitscheiding van de brugfactor MFGE8, die apoptotische cellen en fagocyten aan elkaar koppelt, bevorderen folliculaire dendritische cellen de selectieve verwijdering van afbraakproducten uit de kiemcellen.[13][14]
Voorkomen van auto-immuniteit
Factor Mfge, voornamelijk geproduceerd in lymfoïde weefsels door folliculaire dendritische cellen, staat erom bekend de opname van apoptotische cellen te bevorderen. Een tekort aan deze factor bij muizen leidt tot een toestand die lijkt op systemische lupus erythematodes. Bovendien ontwikkelen muizen zonder lymphotoxine of lymphotoxine-receptoren, die dus geen folliculaire dendritische cel bevatten, gegeneraliseerde lymfocytaire infiltraten, wat wijst op auto-immuniteit. Deze bevindingen suggereren dat folliculaire dendritische cellen mogelijk organismen beschermen tegen auto-immuniteit door de verwijdering van potentieel zelfreactieve afbraakproducten uit kiemcentra.[13]
Interactie met B-cellen
Niet-antigeenspecifieke B-cellen spelen een belangrijke rol bij het transport van antigenen naar folliculaire dendritische cellen. Ze vangen immuuncomplexen op CR1/2-afhankelijke wijze, rechtstreeks uit de lymfe of uit macrofagen en verplaatsen zich naar het lymfeweefsel, waar ze complement-geopsoniseerd antigeen overdragen aan de folliculaire dendritische cellen.[15][16]
Folliculaire dendritische cellen trekken op hun beurt B-cellen aan met chemo-attractant CXCL13. B-cellen zonder CXCR5, de receptor voor CXCL13, dringen nog steeds de witte pulpa binnen, maar zijn verkeerd gelokaliseerd en ongeorganiseerd. Om folliculaire structuren te genereren, moeten folliculaire dendritische cellen worden gestimuleerd door lymfotoxine (LT), een mediator die door B-cellen wordt geproduceerd. De stimulatie van CXCR5 op B-cellen verhoogt de LT-productie, wat leidt tot activering van folliculaire dendritische cellen en verdere CXCL13-secretie stimuleert, waardoor een positieve voorwaartskoppelinglus ontstaat. Dit resulteert in de vorming van kiemcentra (GC's), waar antigeen-geactiveerde B-cellen worden gevangen om somatische mutatie, positieve en negatieve selectie, immunoglobuline class switching en differentiatie te ondergaan in plasmacellen met hoge affiniteit en geheugen-B-cellen.Celadhesie tussen folliculaire dendritische cellen en B-cellen wordt gemedieerd door ICAM-1 (CD54)–LFA-1 (CD11a) en VCAM–VLA-4 moleculen.[7] Geactiveerde B-cellen met een lage affiniteit voor antigeen dat op het oppervlak van folliculaire dendritische cellen is gevangen, evenals autoreactieve B-cellen, ondergaan apoptose,[17] terwijl B-cellen gebonden aan folliculaire dendritische cellen via het antigeencomplex, overleven vanwege apoptoseblokkering veroorzaakt door interactie met folliculaire dendritische cellen.
Klinische betekenis
Het folliculair dendritisch celsarcoom is een uiterst zeldzame neoplasie. Hoewel het bestaan van folliculaire dendritische cel-tumoren al in 1978 door Lennert werd voorspeld, werd de tumor pas in 1986 volledig als een aparte kanker herkend na karakterisering door Monda et al.[18][19] Het vertegenwoordigt slechts 0,4% van de wekedelensarcomen, maar heeft een aanzienlijk recidief- en metastasenpotentieel.
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Follicular dendritic cells op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
- ↑ Liu Y, Grouard G, de Bouteiller O, Banchereau J (1996). Follicular Dendritic Cells and Germinal Centers. DOI:10.1016/S0074-7696(08)62508-5, 139–79. ISBN 978-0-12-364570-8.
- ↑ Heesters, Balthasar A., Myers, Riley C., Carroll, Michael C. (20 juni 2014). Follicular dendritic cells: dynamic antigen libraries. Nature Reviews Immunology 14 (7): 495–504. ISSN: 1474-1733. PMID 24948364. DOI: 10.1038/nri3689.
- ↑ Aguzzi, Adriano, Kranich, Jan, Krautler, Nike Julia (March 2014). Follicular dendritic cells: origin, phenotype, and function in health and disease. Trends in Immunology 35 (3): 105–113. ISSN: 1471-4906. PMID 24315719. DOI: 10.1016/j.it.2013.11.001.
- ↑ Banchereau J, Steinman RM (1998). Dendritic cells and the control of immunity. Nature 392 (6673): 245–52. PMID 9521319. DOI: 10.1038/32588. van Nierop K, de Groot C (2002). Human follicular dendritic cells: function, origin and development. Semin Immunol 14 (4): 251–7. PMID 12163300. DOI: 10.1016/S1044-5323(02)00057-X.
- 1 2 Male D, Brostoff J, Roth D, Roitt I (2007). Immunology, 7th. Elsevier Health Sciences. ISBN 978-0-323-03399-2.
- ↑ Banchereau J, Steinman RM (1998). Dendritic cells and the control of immunity. Nature 392 (6673): 245–52. PMID 9521319. DOI: 10.1038/32588.
- 1 2 3 van Nierop K, de Groot C (2002). Human follicular dendritic cells: function, origin and development. Semin Immunol 14 (4): 251–7. PMID 12163300. DOI: 10.1016/S1044-5323(02)00057-X.
- ↑ Kapasi ZF, Qin D, Kerr WG, Kosco-Vilbois MH, Shultz LD, Tew JG, Szakal AK (1998). Follicular dendritic cell (FDC) precursors in primary lymphoid tissues. The Journal of Immunology 160 (3): 1078–84. PMID 9570519. DOI: 10.4049/jimmunol.160.3.1078.
- ↑ Wang Y, Wang J, Sun Y, Wu Q, Fu YX (2001). Complementary effects of TNF and lymphotoxin on the formation of germinal center and follicular dendritic cells. Journal of Immunology 166 (1): 330–7. PMID 11123309. DOI: 10.4049/jimmunol.166.1.330.
- ↑ Ettinger R, Mebius R, Browning JL, Michie SA, van Tuijl S, Kraal G, van Ewijk W, McDevitt HO (1998). Effects of tumor necrosis factor and lymphotoxin on peripheral lymphoid tissue development. Int Immunol 10 (6): 727–41. PMID 9678753. DOI: 10.1093/intimm/10.6.727.
- ↑ Cyster JG (2010). B cell follicles and antigen encounters of the third kind.. Nat Immunol 11 (11): 989–96. PMID 20959804. DOI: 10.1038/ni.1946.
- ↑ Balthasar, Heesters, Priyadarshini, Chatterjee, Young-A, Kim, Santiago, Gonzalez, Michael, Kuligowski (2013). Endocytosis and recycling of immune complexes by follicular dendritic cells enhances B cell binding and activation. Frontiers in Immunology 4. ISSN: 1664-3224. DOI: 10.3389/conf.fimmu.2013.02.00438.
- 1 2 Aguzzi A, Krautler NJ (2010). Characterizing follicular dendritic cells: A progress report.. European Journal of Immunology 40 (8): 2134–8. PMID 20853499. DOI: 10.1002/eji.201040765.
- ↑ Kranich J, Krautler NJ, Heinen E, Polymenidou M, Bridel C, Schildknecht A, Huber C, Kosco-Vilbois MH, Zinkernagel R, Miele G, Aguzzi A (2008). Follicular dendritic cells control engulfment of apoptotic bodies by secreting Mfge8.. J Exp Med 205 (6): 1293–302. PMID 18490487. PMC 2413028. DOI: 10.1084/jem.20071019.
- ↑ Phan, Tri Giang, Grigorova, Irina, Okada, Takaharu, Cyster, Jason G (29 juli 2007). Subcapsular encounter and complement-dependent transport of immune complexes by lymph node B cells. Nature Immunology 8 (9): 992–1000. ISSN: 1529-2908. PMID 17660822. DOI: 10.1038/ni1494.
- ↑ Carrasco, Yolanda R., Batista, Facundo D. (July 2007). B Cells Acquire Particulate Antigen in a Macrophage-Rich Area at the Boundary between the Follicle and the Subcapsular Sinus of the Lymph Node. Immunity 27 (1): 160–171. ISSN: 1074-7613. PMID 17658276. DOI: 10.1016/j.immuni.2007.06.007.
- ↑ Aguzzi A, Kranich J, Krautler NJ (2014). Follicular dendritic cells: origin, phenotype, and function in health and disease. Trends in Immunology 35 (3): 105–113. PMID 24315719. DOI: 10.1016/j.it.2013.11.001.
- ↑ Lennert, Karl (1978). Malignant lymphomas other than Hodgkin's disease, histology, cytology, ultrastructure, immunology. Springer-Verlag, Berlin, 59–64. ISBN 978-0-387-08020-8.
- ↑ Monda, Lauren, Warnke, Roger, Rosai, Juan (1986). A primary lymph node malignancy with features suggestive of dendritic reticulum cell differentiation. A report of 4 cases. The American Journal of Pathology 122 (3): 562–72. PMID 2420185. PMC 1888214.