Embryonale stamcellen

Embryonale stamcellen (ESC) zijn pluripotente stamcellen die afkomstig zijn uit een zeer vroeg stadium van het embryo, meestal het blastocyststadium (ongeveer 4-5 dagen na de bevruchting bij mensen). Een belangrijk kenmerk van deze cellen is dat ze pluripotent zijn: ze kunnen zich ontwikkelen tot alle celtypen van het lichaam, zoals zenuwcellen, spiercellen en bloedcellen.
Door deze bijzondere eigenschappen zijn embryonale stamcellen van groot belang in wetenschappelijk onderzoek. Ze worden gebruikt bij het bestuderen van de vroege menselijke ontwikkeling, bijvoorbeeld om te begrijpen welke transcriptiefactoren en signaalroutes de eerste celbeslissingen aansturen. Daarnaast worden ze gebruikt bij de ontwikkeling van nieuwe therapieën (regeneratieve geneeskunde) en het testen van medicijnen op ESC-afgeleide cellen, waardoor dierproeven deels overbodig worden.
Het gebruik van embryonale stamcellen voor onderzoek werpt ethische vragen op, omdat voor het verkrijgen van deze cellen een embryo wordt vernietigd. Daarom is er ook veel onderzoek naar alternatieven, zoals geïnduceerde pluripotente stamcellen (iPSC), die uit volwassen lichaamscellen worden gemaakt maar vergelijkbare eigenschappen hebben. In Nederland en België zijn wettelijke kaders vastgesteld voor biomedisch onderzoek met embryonaal materiaal.
Eigenschappen
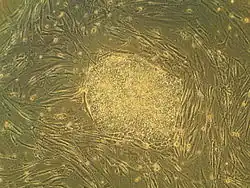
Embryonale stamcellen (ESC's), afkomstig uit het blastocyststadium van vroege zoogdierembryo's, onderscheiden zich door hun vermogen om te differentiëren tot elk embryonaal celtype en door hun vermogen om zichzelf te vernieuwen. Het zijn deze eigenschappen die ze waardevol maken op wetenschappelijk en medisch gebied. ESC's hebben een normaal karyotype, behouden een hoge telomerase-activiteit en vertonen een opmerkelijk proliferatief potentieel op de lange termijn.[1]
Pluripotent
Embryonale stamcellen van de embryoblast zijn pluripotent, wat betekent dat ze kunnen differentiëren om primitief ectoderm te genereren, dat uiteindelijk tijdens de gastrulatie differentieert in alle derivaten van de drie primaire kiembladen: ectoderm, endoderm en mesoderm. Deze kiembladen genereren elk van de meer dan 400 celtypen in het volwassen menselijk lichaam. Wanneer ze de juiste signalen krijgen, vormen ESC's aanvankelijk voorlopercellen die vervolgens differentiëren tot de gewenste celtypen. Pluripotentie onderscheidt embryonale stamcellen van volwassen stamcellen, die multipotent zijn en slechts een beperkt aantal celtypen kunnen produceren.
Zelfvernieuwing en structuurherstel
Onder bepaalde omstandigheden kunnen embryonale stamcellen in een ongedifferentieerde staat zichzelf onbeperkt te vernieuwen. Zelfvernieuwingsomstandigheden moeten voorkomen dat de cellen klonteren en een omgeving in stand houden die een niet-gespecialiseerde staat ondersteunt.[2] Meestal gebeurt dit in het laboratorium met media die serum en leukemie-remmende factor bevatten of serumvrije mediasupplementen met twee remmende geneesmiddelen ("2i"), de MEK-remmer PD03259010 en GSK-3-remmer CHIR99021.[3]
Groei
ESC's delen zich zeer frequent vanwege een verkorte G1-fase in hun celcyclus. Snelle celdeling zorgt ervoor dat de cellen snel in aantal toenemen, maar niet in grootte, wat belangrijk is voor de vroege embryonale ontwikkeling. In ESC's worden cycline A- en cycline E-eiwitten die betrokken zijn bij de G1/S-overgang altijd op hoge niveaus tot expressie gebracht.[4] Cycline-afhankelijke kinases zoals CDK2 die de voortgang van de celcyclus bevorderen, zijn overactief, deels vanwege de downregulatie van hun remmers.[5] Retinoblastoom-eiwitten die de transcriptiefactor E2F remmen totdat de cel klaar is om de S-fase in te gaan, worden in ESC's hypergefosforyleerd en geïnactiveerd, wat leidt tot voortdurende expressie van proliferatiegenen.[4] Deze veranderingen resulteren in versnelde cycli van celdeling. Hoewel hoge expressieniveaus van pro-proliferatieve eiwitten en een verkorte G1-fase in verband zijn gebracht met het behoud van pluripotentie,[6][7] brengen ESC's die gekweekt worden in serumvrije 2i-omstandigheden wel hypo-gefosforyleerde actieve retinoblastoom-eiwitten tot expressie en hebben ze een verlengde G1-fase.[8] Ondanks dit verschil in de celcyclus in vergelijking met ESC's die gekweekt worden in media met serum, hebben deze cellen vergelijkbare pluripotente kenmerken.[9] Pluripotentiefactoren Oct4 en Nanog spelen een rol bij de transcriptionele regulering van de embryonale stamcelcyclus.[10][11]
Toepassingen
Embryonale stamcellen kunnen zich in vrijwel alle mogelijke celtypen ontwikkelen en zich in principe onbeperkt vernieuwen. Daarom worden ze onderzocht als mogelijk toepassing in de regeneratieve geneeskunde, bijvoorbeeld als mogelijke bron van donorweefsel. ESC's zijn onder meer gebruikt bij aandoeningen als ruggenmergletsel, diabetes en neurodegeneratieve ziekten. Buiten therapie worden ESC’s ook gebruikt om de vroege menselijke ontwikkeling in kaart te brengen, en genetische ziekten of toxische effecten van stoffen in het laboratorium te bestuderen.
Weefseltechnologie
In de weefseltechnologie is het gebruik van stamcellen van belang. Om een weefsel succesvol te kunnen ontwerpen, moeten de gebruikte cellen specifieke biologische functies kunnen uitvoeren, zoals de secretie van cytokines, signaalmoleculen, interactie met naburige cellen en het produceren van een extracellulaire matrix in de juiste organisatie. Stamcellen vertonen deze specifieke biologische functies, samen met het vermogen om zichzelf te vernieuwen en te differentiëren in een of meer typen gespecialiseerde cellen. Embryonale stamcellen zijn een van de bronnen die worden overwogen voor het gebruik in de weefseltechnologie. Het gebruik van menselijke embryonale stamcellen heeft veel nieuwe mogelijkheden geopend voor de weefseltechnologie, maar er zijn nog veel horden die moeten worden genomen voordat menselijke embryonale stamcellen zelfs maar kunnen worden gebruikt. Er wordt verondersteld dat als embryonale stamcellen zodanig kunnen worden aangepast dat ze geen immuunreactie meer oproepen wanneer ze bij de patiënt worden geïmplanteerd, dit een revolutionaire stap zou zijn in de weefseltechnologie.[12] Embryonale stamcellen zijn niet beperkt tot de weefseltechnologie.
Celvervangende therapieën
Onderzoek heeft zich gericht op het differentiëren van ESC's in verschillende celtypen voor uiteindelijk gebruik als celvervangende therapieën. Enkele van de celtypen die zijn of momenteel worden ontwikkeld, zijn hartspiercellen, zenuwcellen, hepatocyten, beenmergcellen, eilandjes van Langerhanscellen en endotheelcellen.[13] De vorming van dergelijke celtypen uit ESC's is echter niet zonder obstakels, daarom heeft het onderzoek zich gericht op het overwinnen van deze obstakels. Er zijn bijvoorbeeld studies gaande om ESC's te differentiëren in weefselspecifieke hartspiercellen en om hun onvolwassen eigenschappen te verwijderen die hen onderscheiden van volwassen hartspiercellen.[14]
Klinisch potentieel
.jpg)
- Onderzoekers hebben ESC's gedifferentieerd in dopamine-producerende cellen in de hoop dat deze zenuwcellen gebruikt kunnen worden bij de behandeling van de ziekte van Parkinson.[15][16]
- ESC's zijn gedifferentieerd in naturalkillercellen en botweefsel.[17]
- Er zijn studies gaande met ESC's om een alternatieve behandeling voor diabetes te bieden. ESC's zijn bijvoorbeeld gedifferentieerd in insuline-producerende cellen,[18] en onderzoekers aan de Harvard University waren in staat om grote hoeveelheden alvleesklier-bètacellen te produceren uit ESC's.[19]
- Een artikel gepubliceerd in het European Heart Journal beschrijft een translationeel proces van het genereren van uit menselijke embryonale stamcellen afgeleide hartvoorlopercellen die gebruikt kunnen worden in klinische proeven met patiënten met ernstig hartfalen.[20]
- Programmeren van voorlopercellen. De ontwikkeling van een protocol voor de gerichte generatie van alvleeskliervoorlopercellen is uitgevoerd op hESC's (menselijke embryonale stamcellen). Deze cellen, die een enorm potentieel hebben in de behandeling van metabole aandoeningen van de alvleesklier zoals diabetes, zijn geprogrammeerd tot alvleeskliervoorlopercellen door het nabootsen van ontwikkelingssignalen, die een zich ontwikkelend endoderm nodig zou hebben om functioneel alvleesklierweefsel te vormen.[21] hESC's worden gekweekt op matrigel en vervolgens gedifferentieerd tot endoderm en verder onder invloed van bFGF, EGF, BMP4.[22]
Onderzoek nieuwe geneesmiddelen
Naast het feit dat ze een belangrijk alternatief zijn voor orgaantransplantaties, worden ESC's ook gebruikt op het gebied van toxicologie en als cellulaire screenings om nieuwe chemische stoffen te ontdekken die kunnen worden ontwikkeld als kleine-moleculaire geneesmiddelen. Studies hebben aangetoond dat hartspiercellen die zijn afgeleid van ESC's gevalideerde in vitro-modellen zijn voor het testen van medicijnreacties en het voorspellen van toxiciteitsprofielen.[13] Van ESC afgeleide hartspiercellen is aangetoond dat ze reageren op farmacologische stimuli en kunnen daarom worden gebruikt om cardiotoxiciteit te beoordelen, zodat ze bijvoorbeeld geen Torsade de Pointes veroorzaken.[23]
ESC-afgeleide hepatocyten zijn ook nuttige modellen die kunnen worden gebruikt in de preklinische stadia van onderzoek naar nieuwe geneesmiddelen. De ontwikkeling van hepatocyten uit ESC's is echter moeilijk gebleken en dit belemmert het vermogen om het metabolisme van geneesmiddelen te testen. Daarom heeft het onderzoek zich gericht op het ontwikkelen van volledig functionele ESC-afgeleide hepatocyten met stabiele fase I- en II-enzymactiviteit.[24]
Modelleren van genetische stoornissen
Verschillende nieuwe studies zijn begonnen met het concept van het modelleren van genetische stoornissen met embryonale stamcellen. Ofwel door genetische manipulatie van de cellen, of meer recent, door het afleiden van zieke cellijnen die zijn geïdentificeerd door prenatale genetische diagnose (PGD), is het modelleren van genetische aandoeningen iets dat is bereikt met stamcellen. Deze aanpak kan heel waardevol blijken bij het bestuderen van aandoeningen zoals het fragiele-X-syndroom, cystische fibrose en andere erfelijke aandoeningen waarvoor geen betrouwbaar modelsysteem bestaat.
Yury Verlinsky, een Russisch-Amerikaanse medisch onderzoeker die zich specialiseerde in embryo- en cellulaire genetica (genetische cytologie), ontwikkelde prenatale diagnostische testmethoden om op genen beruste en chromosomale aandoeningen anderhalve maand eerder te bepalen dan met een standaard vruchtwaterpunctie. De technieken worden nu gebruikt door veel zwangere vrouwen en toekomstige ouders, met name stellen met een voorgeschiedenis van genetische aandoemingen of waarbij de vrouw ouder is dan 35 (wanneer het risico op genetisch gerelateerde aandoeningen hoger is). Bovendien hebben ouders, door een embryo zonder genetische aandoeningen te selecteren, de mogelijkheid om het leven van broers en zussen te redden die al soortgelijke aandoeningen en ziekten hebben met behulp van cellen van het ziektevrije nageslacht. Dit wordt een savior baby, een donorbaby, een reddingsbaby of een baby op maat genoemd. Hierbij wordt in-vitrofertilisatie gebruikt, waarbij de bevruchte eicel hetzelfde genetische humaan leukocytenantigeen (HLA-type) van een broertje of zusje met een bepaalde ziekte heeft. De bevruchte eicel is dus zelf vrij van de ziekte én heeft een kans om mesenchymatische stamcellen uit de navelstreng met stamceltransplantatie door te geven voor het genezen van de broer of zus, die lijdt aan een dodelijke ziekte, zoals kanker of anemie van Fanconi, die het beste behandeld kunnen worden met een hematopoëtische stamceltransplantatie. Bevruchte eicellen worden getest op genetische compatibiliteit (typering van (HLA)), met behulp van pre-implantatiegenetische diagnostiek (PGD), en alleen bevruchte eicellen die compatibel zijn met het bestaande kind worden geïmplanteerd. Bevruchte eicellen worden ook getest om er zeker van te zijn dat ze vrij zijn van de oorspronkelijke genetische ziekte. De procedure is echter controversieel.[25]
Herstel van DNA-schade

Gedifferentieerde somatische cellen en embryonale stamcellen (ES-cellen) gebruiken verschillende strategieën om met DNA-schade om te gaan. Bijvoorbeeld, menselijke voorhuidfibroblasten, een type somatische cel, gebruiken niet-homologe eindbinding (NHEJ), een foutgevoelig DNA-herstelproces, als het primaire proces voor het repareren van dubbelstrengsbreuken tijdens alle stadia van de celcyclus.[26] Vanwege zijn foutgevoelige aard, heeft NHEJ de neiging om mutaties te produceren in de klonale afstammelingen van een cel.
Embryonale stamcellen (ES) gebruiken een andere strategie om met dubbelstrengsbreuken om te gaan.[27] Omdat ES-cellen ten grondslag liggen van alle celtypen van een organisme, inclusief de cellen van de kiembaan, zijn mutaties die ontstaan in ES-cellen als gevolg van gebrekkige DNA-reparatie een ernstiger probleem dan in gedifferentieerde somatische cellen. Bijgevolg zijn robuuste mechanismen nodig in ES-cellen om DNA-schade nauwkeurig te repareren, en als reparatie mislukt, om die cellen met niet-gerepareerde DNA-schade te verwijderen. Zo gebruiken muizen-ES-cellen voornamelijk homologe recombinatiereparatie (HRR) om dubbelstrengsbreuken te repareren.[27] Dit type reparatie is afhankelijk van de interactie van de twee zusterchromatiden die tijdens de S-fase worden gevormd en samen aanwezig zijn tijdens de G2-fase van de celcyclus. HRR kan dubbelstrengsbreuken in één zusterchromatide nauwkeurig repareren door intacte informatie van het andere zusterchromatide te gebruiken. Cellen in de G1-fase van de celcyclus (d.w.z. na metafase/celdeling maar vóór de volgende ronde van replicatie) hebben slechts één kopie van elk chromosoom (d.w.z. zusterchromatiden zijn niet aanwezig). Muizen-ES-cellen missen een G1-controlepunt en stoppen de celcyclus niet bij het oplopen van DNA-schade.[28] In plaats daarvan ondergaan ze geprogrammeerde celdood (apoptose) als reactie op DNA-schade.[29] Apoptose kan worden gebruikt als een failsafe strategie om cellen met niet-gerepareerde DNA-schade te verwijderen om mutatie en progressie naar kanker te voorkomen.[30] In overeenstemming met deze strategie hebben muizen-embryonale stamcellen een mutatiefrequentie die ongeveer 100 maal lager is dan die van klonale somatische cellen.[31]
Zie ook
- ↑ Thomson, J. A., Itskovitz-Eldor, J, Shapiro, S. S., Waknitz, M. A., Swiergiel, J. J. (1998). Embryonic Stem Cell Lines Derived from Human Blastocysts. Science 282 (5391): 1145–7. PMID 9804556. DOI: 10.1126/science.282.5391.1145.
- ↑ Ying (2003). BMP Induction of Id Proteins Suppresses Differentiation and Sustains Embryonic Stem Cell Self-Renewal in Collaboration with STAT3. Cell 115 (3): 281–292. PMID 14636556. DOI: 10.1016/S0092-8674(03)00847-X.
- ↑ Martello, G., Smith, A. (2014). The nature of embryonic stem cells. Annual Review of Cell and Developmental Biology 30: 647–75. PMID 25288119. DOI: 10.1146/annurev-cellbio-100913-013116.
- 1 2 Boward, B., Wu, T., Dalton, S. (2016). Concise Review: Control of Cell Fate Through Cell Cycle and Pluripotency Networks. Stem Cells 34 (6): 1427–36. PMID 26889666. PMC 5201256. DOI: 10.1002/stem.2345.
- ↑ White, J., Stead, E., Faast, R., Conn, S., Cartwright, P. (2005). Developmental activation of the Rb-E2F pathway and establishment of cell cycle-regulated cyclin-dependent kinase activity during embryonic stem cell differentiation. Molecular Biology of the Cell 16 (4): 2018–27. PMID 15703208. PMC 1073679. DOI: 10.1091/mbc.e04-12-1056.
- ↑ Ter Huurne, Menno, Stunnenberg, Hendrik G. (21 april 2021). G1-phase progression in pluripotent stem cells. Cellular and Molecular Life Sciences 21 (10): 4507–4519. ISSN: 1875-9777. PMID 33884444. PMC 8195903. DOI: 10.1007/s00018-021-03797-8.
- ↑ Singh, Amar M., Dalton, Stephen (7 augustus 2009). The cell cycle and Myc intersect with mechanisms that regulate pluripotency and reprogramming. Cell Stem Cell 5 (2): 141–149. ISSN: 1875-9777. PMID 19664987. PMC 2909475. DOI: 10.1016/j.stem.2009.07.003.
- ↑ Ter Huurne, Menno, Chappell, James, Dalton, Stephen, Stunnenberg, Hendrik G. (5 October 2017). Distinct Cell-Cycle Control in Two Different States of Mouse Pluripotency. Cell Stem Cell 21 (4): 449–455.e4. ISSN: 1875-9777. PMID 28985526. PMC 5658514. DOI: 10.1016/j.stem.2017.09.004.
- ↑ Ying, Qi-Long, Wray, Jason, Nichols, Jennifer, Batlle-Morera, Laura, Doble, Bradley (22 mei 2008). The ground state of embryonic stem cell self-renewal. Nature 453 (7194): 519–523. ISSN: 1476-4687. PMID 18497825. PMC 5328678. DOI: 10.1038/nature06968.
- ↑ Lee, J., Go, Y., Kang, I., Han, Y. M., Kim, J. (2010). Oct-4 controls cell-cycle progression of embryonic stem cells. The Biochemical Journal 426 (2): 171–81. PMID 19968627. PMC 2825734. DOI: 10.1042/BJ20091439.
- ↑ Zhang, X., Neganova, I., Przyborski, S., Yang, C., Cooke, M. (2009). A role for NANOG in G1 to S transition in human embryonic stem cells through direct binding of CDK6 and CDC25A. The Journal of Cell Biology 184 (1): 67–82. PMID 19139263. PMC 2615089. DOI: 10.1083/jcb.200801009.
- ↑ (en) Heath, Carole A. (1 januari 2000). Cells for tissue engineering. Trends in Biotechnology 18 (1): 17–19. ISSN: 0167-7799. PMID 10631775. DOI: 10.1016/S0167-7799(99)01396-7. Gearchiveerd op 13 april 2021. Geraadpleegd op 13 april 2021.
- 1 2 Davila, JC, Cezar, GG, Thiede, M, Strom, S, Miki, T (2004). Use and application of stem cells in toxicology. Toxicological Sciences 79 (2): 214–223. PMID 15014205. DOI: 10.1093/toxsci/kfh100.
- ↑ Siu, CW, Moore, JC, Li, RA (2007). Human embryonic stem cell-derived cardiomyocytes for heart therapies. Cardiovascular & Hematological Disorders Drug Targets 7 (2): 145–152. PMID 17584049. DOI: 10.2174/187152907780830851.
- ↑ Perrier, A. L. (2004). Derivation of midbrain dopamine neurons from human embryonic stem cells. Proceedings of the National Academy of Sciences 101 (34): 12543–12548. PMID 15310843. PMC 515094. DOI: 10.1073/pnas.0404700101.
- ↑ Parish, CL, Arenas, E (2007). Stem-cell-based strategies for the treatment of Parkinson's disease. Neuro-Degenerative Diseases 4 (4): 339–347. PMID 17627139. DOI: 10.1159/000101892.
- ↑ Waese, EY, Kandel, RA, Stanford, WL (2008). Application of stem cells in bone repair. Skeletal Radiology 37 (7): 601–608. PMID 18193216. DOI: 10.1007/s00256-007-0438-8.
- ↑ d'Amour, KA, Bang, AG, Eliazer, S, Kelly, OG, Agulnick, AD (2006). Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nature Biotechnology 24 (11): 1392–1401. PMID 17053790. DOI: 10.1038/nbt1259.
- ↑ Colen, B.D. (9 October 2014) Giant leap against diabetes Gearchiveerd 2014-12-02 in de Wayback Machine. The Harvard Gazette, Retrieved 24 November 2014
- ↑ Menasché, Phillip, Vanneaux, Valérie, Fabreguettes, Jean-Roch, Bel, Alain, Tosca, Lucie (21 March 2015). Towards a clinical use of human embryonic stem cell derived-cardiac progenitors: a translational experience. European Heart Journal 36 (12): 743–750. PMID 24835485. DOI: 10.1093/eurheartj/ehu192.
- ↑ Chen, S., Borowiak, M., Fox, J. L., Maehr, R., Osafune, K. (2009). A small molecule that directs differentiation of human ESCs into the pancreatic lineage. Nature Chemical Biology 5 (4): 258–265. PMID 19287398. DOI: 10.1038/nchembio.154.
- ↑ Zaret, K. S. (2008). Genetic programming of liver and pancreas progenitors: lessons for stem-cell differentiation. Nature Reviews Genetics 9 (5): 329–340. PMID 18398419. DOI: 10.1038/nrg2318.
- ↑ Jensen, J, Hyllner, J, Björquist, P (2009). Human embryonic stem cell technologies and drug discovery. Journal of Cellular Physiology 219 (3): 513–519. PMID 19277978. DOI: 10.1002/jcp.21732.
- ↑ Söderdahl, T, Küppers-Munther, B, Heins, N, Edsbagge, J, Björquist, P (2007). Glutathione transferases in hepatocyte-like cells derived from human embryonic stem cells. Toxicology in Vitro 21 (5): 929–937. PMID 17346923. DOI: 10.1016/j.tiv.2007.01.021.
- ↑ Wat is een "savior baby"?
- ↑ Mao Z, Bozzella M, Seluanov A, Gorbunova V (September 2008). DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells. Cell Cycle 7 (18): 2902–2906. PMID 18769152. PMC 2754209. DOI: 10.4161/cc.7.18.6679.
- 1 2 Tichy ED, Pillai R, Deng L (November 2010). Mouse embryonic stem cells, but not somatic cells, predominantly use homologous recombination to repair double-strand DNA breaks. Stem Cells Dev. 19 (11): 1699–1711. PMID 20446816. PMC 3128311. DOI: 10.1089/scd.2010.0058.
- ↑ Hong Y, Stambrook PJ (October 2004). Restoration of an absent G1 arrest and protection from apoptosis in embryonic stem cells after ionizing radiation. Proc. Natl. Acad. Sci. U.S.A. 101 (40): 14443–14448. PMID 15452351. PMC 521944. DOI: 10.1073/pnas.0401346101.
- ↑ Aladjem MI, Spike BT, Rodewald LW (January 1998). ES cells do not activate p53-dependent stress responses and undergo p53-independent apoptosis in response to DNA damage. Curr. Biol. 8 (3): 145–155. PMID 9443911. DOI: 10.1016/S0960-9822(98)70061-2.
- ↑ Bernstein C, Bernstein H, Payne CM, Garewal H (June 2002). DNA repair/pro-apoptotic dual-role proteins in five major DNA repair pathways: fail-safe protection against carcinogenesis. Mutat. Res. 511 (2): 145–178. PMID 12052432. DOI: 10.1016/S1383-5742(02)00009-1.
- ↑ Cervantes RB, Stringer JR, Shao C, Tischfield JA, Stambrook PJ (March 2002). Embryonic stem cells and somatic cells differ in mutation frequency and type. Proc. Natl. Acad. Sci. U.S.A. 99 (6): 3586–3590. PMID 11891338. PMC 122567. DOI: 10.1073/pnas.062527199.