Retinale horizontale cel
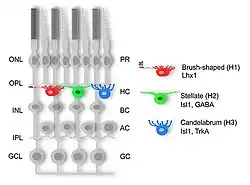
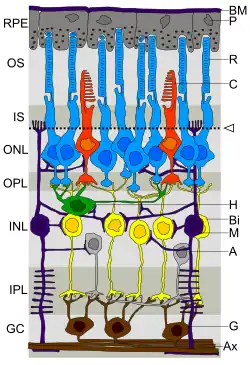

Retinale horizontale cellen zijn de lateraal met elkaar verbonden zenuwcellen met cellichamen in de binnenste nucleaire laag van het netvlies van gewervelden. Ze ontstaan uit multipotente retinale voorlopercellen in de oogbeker van gewervelden[1] en helpen bij het integreren en reguleren van de input van meerdere lichtgevoelige cellen. Men neemt aan dat retinale horizontale cellen onder andere verantwoordelijk zijn voor het verhogen van het contrast via laterale inhibitie en voor de aanpassing aan zowel lichte als donkere lichtomstandigheden. Retinale horizontale cellen zorgen voor remmende terugkoppeling naar staafjes- en kegeltjes.[2][3] Men denkt dat ze belangrijk zijn voor de antagonistische centrum-surround-eigenschap van de receptieve velden van veel soorten retinale ganglioncellen.[4]
Retinale horizontale cellen kunnen worden onderverdeeld in axondragende en axonloze subtypen, een kenmerk dat behouden lijkt te blijven in de lijn van gewervelden, op enkele uitzonderingen na. Bij netvliezen van gewervelden is de retinale horizontale cel met een axon universeel. Subtypen hebben geen axon. In het kippennetvlies zijn drie verschillende subtypen beschreven op basis van morfologie; het "borstelvormige" (H1) met een axon, terwijl het "stellaatvormige" (H2) en het "kandelabervormige" (H3) beide gen axon hebben. Bij sommige soorten worden deze subtypen aangeduid als A-, B- en C-type. Deze morfologische classificatie correleert met de expressie van unieke moleculaire markers.[5]
Structuur
Afhankelijk van de soort zijn er doorgaans één of twee klassen horizontale cellen, waarbij soms een derde type wordt voorgesteld.[2][3]
Retinale horizontale cellen strekken zich uit over lichtgevoelige cellen en tellen de input op voordat ze synapsen vormen met lichtgevoelige cellen.[2][3] Retinale horizontale cellen zouden ook synapsen kunnen vormen met retinale bipolaire cellen, maar dit is nog onzeker.[2][3] Horizontal cells may also synapse onto bipolar cells, but this remains uncertain.[2][6]
De dichtheid van retinale horizontale cellen is groter richting het centrale deel van het netvlies. Bij de kat is waargenomen dat A-type retinale horizontale cellen een dichtheid hebben van 225 cellen/mm² nabij het midden van het netvlies en een dichtheid van 120 cellen/mm² in de meer perifere retina.[7]
De kans dat retinale horizontale cellen en andere interneuronen in het netvlies zich dicht bij elkaar bevinden, is kleiner dan volgens toeval, wat resulteert in 'uitsluitingszones' die ze scheiden. Mozaïekrangschikkingen bieden een mechanisme om elk celtype gelijkmatig over het netvlies te verdelen, waardoor alle delen van het gezichtsveld toegang hebben tot een volledige set verwerkingselementen.[7] De transmembraaneiwitten MEGF10 en MEGF11 spelen een cruciale rol bij de vorming van de mozaïeken door retinale horizontale cellen en starburst-amacriene cellen bij muizen.[8]
Functie
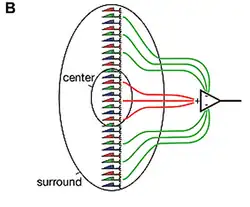
Retinale horizontale cellen worden gedepolariseerd door de afgifte van glutamaat uit lichtgevoelige cellen, wat gebeurt bij afwezigheid van licht. Depolarisatie van een retinale horizontale cel zorgt ervoor dat nabijgelegen lichtgevoelige cellen hyperpolariseren. Omgekeerd geeft een lichtgevoelige cel in het licht minder glutamaat af, wat de retinale horizontale cel hyperpolariseert, wat leidt tot depolarisatie van nabijgelegen lichtgevoelige cellen. Retinale horizontale cellen zorgen dus voor negatieve terugkoppeling aan lichtgevoelige cellen. De matig brede laterale spreiding en koppeling van retinale horizontale cellen door gap junctions, meet het gemiddelde lichtniveau dat op een gebied van het netvliesoppervlak valt. Deze retinale horizontale cellen trekken vervolgens een evenredige waarde af van de output van lichtgevoelige cellen om de signaalinvoer naar het interne netvliescircuit binnen het werkingsbereik te houden.[2] Retinale horizontale cellen zijn ook een van de twee groepen remmende interneuronen die bijdragen aan de omgeving (surround) van retinale ganglioncellen.[3]
Belichting Centrum lichtgevoelige cel hyperpolarisatie Retinale horizontale cel hyperpolarisatie Surround lichtgevoelige cel depolarisatie

Het exacte mechanisme waarmee depolarisatie van retinale horizontale cellen lichtgevoelige cellen hyperpolariseert is onduidelijk. Hoewel retinale horizontale cellen GABA bevatten, omvatten de belangrijkste mechanismen waarmee retinale horizontale cellen kegeltjes remmen waarschijnlijk niet de afgifte van GABA door retinale horizontale cellen op kegeltjes..[6][9][10] Twee mechanismen, die elkaar niet uitsluiten, dragen waarschijnlijk bij aan de remming van de retinale horizontale cellen door de glutamaatafgifte van de kegeltjes. Beide veronderstelde mechanismen zijn afhankelijk van de beschermde omgeving die wordt geboden door de invaginerende synapsen die retinale horizontale cellen op kegeltjes maken.[6][11] Het eerste veronderstelde mechanisme is een zeer snel efaptisch mechanisme dat geen synaptische vertraging heeft, waardoor het een van de snelste remmende synapsen is die bekend zijn.[6][12][13] Het tweede veronderstelde mechanisme is relatief langzaam met een tijdconstante van ongeveer 200 ms en is afhankelijk van ATP-afgifte via Pannexine 1-kanalen die zich bevinden op de dendrieten van retinale horizontale cellen die de synaptische terminal van de kegel invagineren. De ecto-ATPase NTPDase1 hydrolyseert extracellulair ATP tot AMP, fosfaatgroepen en protonen. De fosfaatgroepen en protonen vormen een pH-buffer met een pKa van 7,2, waardoor de pH in de synaptische spleet relatief zuur blijft. Dit remt de Ca2+-kanalen in de kegeltjes en vermindert daardoor de glutamaatafgifte door de kegeltjes.[6][13][14][15][16]
Het centrum-surround-antagonisme van retinale bipolaire cellen wordt vermoedelijk geërfd van kegeltjes. Wanneer echter opnames worden gemaakt van delen van het kegeltje die zich ver van de kegeltjesuiteinden bevinden die synapsen met bipolaire cellen vormen, lijkt centrum-surround-antagonisme minder betrouwbaar te zijn in kegeltjes dan in retinale bipolaire cellen. Omdat de invaginerende synapsen van retinale horizontale cellen aan kegeltjesuiteinden worden gevormd, wordt gedacht dat het centrum-surround-antagonisme van kegeltjes betrouwbaarder aanwezig is in kegeltjesuiteinden.[17]
Externe link
- Illustratie afkomstig uit Mills en Massey, 1994 (67) - Her retinale horizontaal celnetwerk van konijnen zichtbaar gemaakt door middel van kleurstofinjectie. Lucifergeel, geïnjecteerd in één cel, verspreidt zich via de gap junctions die retinale horizontale cellen van type A verbinden, waardoor de centraal geïnjecteerde cel en honderden aangrenzende cellen zichtbaar worden.
{kind=link}
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Retina horizontal cell op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
- ↑ Boije H, Shirazi Fard S, Edqvist PH and Hallböök F (2016) Horizontal Cells, the Odd Ones Out in the Retina, Give Insights into Development and Disease. Front. Neuroanat. 10:77. doi: 10.3389/fnana.2016.00077
- 1 2 3 4 5 6 Masland, RH (2012). The neuronal organization of the retina. Neuron 76 (2): 266–280. PMID 23083731. PMC 3714606. DOI: 10.1016/j.neuron.2012.10.002.
- 1 2 3 4 5 Demb JB, Singer JH (November 2015). Functional Circuitry of the Retina. Annu Rev Vis Sci 1: 263–289. PMID 28532365. PMC 5749398. DOI: 10.1146/annurev-vision-082114-035334.
- ↑ (en) Chaya, Taro, Matsumoto, Akihiro, Sugita, Yuko, Watanabe, Satoshi, Kuwahara, Ryusuke (17 juli 2017). Versatile functional roles of horizontal cells in the retinal circuit. Scientific Reports 7 (1). ISSN: 2045-2322. PMID 28717219. PMC 5514144. DOI: 10.1038/s41598-017-05543-2.
- ↑ Boije H, Shirazi Fard S, Edqvist PH and Hallböök F (2016) Horizontal Cells, the Odd Ones Out in the Retina, Give Insights into Development and Disease. Front. Neuroanat. 10:77. doi: 10.3389/fnana.2016.00077
- 1 2 3 4 5 Thoreson WB, Mangel SC (September 2012). Lateral interactions in the outer retina. Prog Retin Eye Res 31 (5): 407–41. PMID 22580106. PMC 3401171. DOI: 10.1016/j.preteyeres.2012.04.003.
- 1 2 Wässle H, Riemann HJ (March 1978). The mosaic of nerve cells in the mammalian retina. Proc. R. Soc. Lond. B Biol. Sci. 200 (1141): 441–61. PMID 26058. DOI: 10.1098/rspb.1978.0026.
- ↑ Kay, JN, Chu, MW, Sanes, JR (2012). MEGF10 and MEGF11 mediate homotypic interactions required for mosaic spacing of retinal neurons. Nature 483 (7390): 465–9. PMID 22407321. PMC 3310952. DOI: 10.1038/nature10877.
- ↑ Verweij J, Kamermans M, Spekreijse H (December 1996). Horizontal cells feed back to cones by shifting the cone calcium-current activation range. Vision Res. 36 (24): 3943–53. PMID 9068848. DOI: 10.1016/S0042-6989(96)00142-3.
- ↑ (November 2003). Surround antagonism in macaque cone photoreceptors. J. Neurosci. 23 (32): 10249–57. PMID 14614083. PMC 6741006. DOI: 10.1523/JNEUROSCI.23-32-10249.2003.
- ↑ Barnes S (Dec 2003). Center-surround antagonism mediated by proton signaling at the cone photoreceptor synapse. J Gen Physiol 122 (6): 653–6. PMID 14610023. PMC 2229589. DOI: 10.1085/jgp.200308947.
- ↑ Kamermans M, Fahrenfort I, Schultz K, Janssen-Bienhold U, Sjoerdsma T, Weiler R (May 2001). Hemichannel-mediated inhibition in the outer retina. Science 292 (5519): 1178–80. PMID 11349152. DOI: 10.1126/science.1060101.
- 1 2 Vroman R, Klaassen LJ, Howlett MH, Cenedese V, Klooster J, Sjoerdsma T, Kamermans M (May 2014). Extracellular ATP hydrolysis inhibits synaptic transmission by increasing ph buffering in the synaptic cleft. PLOS Biol. 12 (5): e1001864. PMID 24844296. PMC 4028192. DOI: 10.1371/journal.pbio.1001864.
- ↑ Hirasawa H, Kaneko A (December 2003). pH changes in the invaginating synaptic cleft mediate feedback from horizontal cells to cone photoreceptors by modulating Ca2+ channels. J. Gen. Physiol. 122 (6): 657–71. PMID 14610018. PMC 2229595. DOI: 10.1085/jgp.200308863.
- ↑ Vessey JP, Stratis AK, Daniels BA, Da Silva N, Jonz MG, Lalonde MR, Baldridge WH, Barnes S (April 2005). Proton-mediated feedback inhibition of presynaptic calcium channels at the cone photoreceptor synapse. J. Neurosci. 25 (16): 4108–17. PMID 15843613. PMC 6724943. DOI: 10.1523/JNEUROSCI.5253-04.2005.
- ↑ Davenport CM, Detwiler PB, Dacey DM (January 2008). Effects of pH buffering on horizontal and ganglion cell light responses in primate retina: evidence for the proton hypothesis of surround formation. J. Neurosci. 28 (2): 456–64. PMID 18184788. PMC 3057190. DOI: 10.1523/JNEUROSCI.2735-07.2008.
- ↑ Byzov AL, Shura-Bura TM (1986). Electrical feedback mechanism in the processing of signals in the outer plexiform layer of the retina. Vision Res. 26 (1): 33–44. PMID 3012877. DOI: 10.1016/0042-6989(86)90069-6.