Retinale bipolaire cel
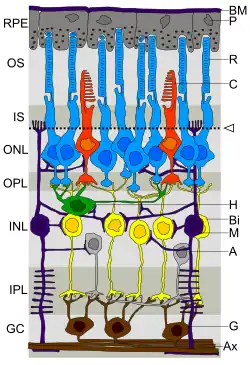

Retinale bipolaire cellen zijn schakelcellen in het netvlies tussen de lichtgevoelige cellen (staafjes en kegeltjes) en retinale ganglioncellen. Ze zorgen, direct of indirect, voor de overdracht van signalen van de lichtgevoelige cellen naar de retinale ganglioncellen.
Structuur
Retinale bipolaire cellen worden zo genoemd omdat ze een centraal lichaam hebben waaruit twee uitlopers ontstaan. Ze kunnen synapsen vormen met staafjes of kegeltjes (gemengde input van staafjes en kegeltjes is aangetroffen bij beenvissen, maar niet bij zoogdieren). De retinale bipolaire cellen geven vervolgens de signalen van de lichtgevoelige cellen door en geven deze direct of indirect (via amacriene cellen) door aan de ganglioncellen. In tegenstelling tot de meeste zenuwcellen communiceren retinale bipolaire cellen via gegradeerde potentialen in plaats van actiepotentialen. Het gegradeerde potentiaal is een verandering in het membraanpotentiaal over een relatief klein deel van het membraan en kunnen lang of kort zijn, afhankelijk van hoe lang de kanalen open blijven in tegenstelling tot actiepotentialen, die een vaste amplitude en tijdsverloop hebben.
Functie

Retinale bipolaire cellen ontvangen synaptische input van staafjes of kegeltjes, of van zowel staafjes als kegeltjes, hoewel ze over het algemeen staafbipolaire of kegeltjesbipolaire cellen worden genoemd. Er zijn 13 verschillende vormen van kegeltjesbipolaire cellen, maar er zijn slechts twee vormen van de staafjesbipolaire cel,[1][2] omdat de staafjesreceptor waarschijnlijk later in de evolutionaire geschiedenis is ontstaan dan de kegeltjesreceptor.[3] Volwassen zebravissen hebben meer dan 20 bipolaire cellen die diverse sets van contacten maken over de vier kegeltypes en één staaftype in het buitenste deel van het netvlies.[4]
In het donker zal een lichtgevoelige cel (staafje/kegeltje) glutamaat afgeven, wat de AAN-bipolaire cellen remt (hyperpolariseert) en de UIT-bipolaire cellen aanstuurt (depolariseert). Onder lichte omstandigheden valt het licht echter op de lichtgevoelige cel, waardoor deze wordt geremd (gehyperpolariseerd) door de activering van opsinen die G-proteïnen activeren die fosfodiësterasen (PDE) activeren, dat cyclisch guanosinemonofosfaat (cGMP) splitst in 5'-GMP. In lichtgevoelige cellen is er een overvloed aan cGMP in het donker, waardoor cGMP-gestuurde natriumkanalen open blijven. Activering van PDE vermindert de cGMP-aanvoer, waardoor het aantal open natriumkanalen afneemt en de lichtgevoelige cel hyperpolariseert, waardoor er minder glutamaat wordt vrijgegeven. Dit zorgt ervoor dat de AAN-bipolaire cel zijn inhibitie verliest en actief wordt (gedepolariseerd), terwijl de UIT-bipolaire cel zijn excitatie (stimulering) verliest (hyperpolariseert) en inactief wordt.[5]
Retinale bipolaire staafjescellen synapsen niet rechtstreeks met ganglioncellen. In plaats daarvan synapsen retinale bipolaire staafjescellen met een retinale amacriene cel, die op zijn beurt AAN-bipolaire kegeltjescellen exciteert (stimuleert) (via gap junctions) en UIT-bipolaire kegeltjescellen remt (via glycine-gemedieerde remmende synapsen), waardoor de kegeltjesbaan wordt overgenomen om signalen naar ganglioncellen te sturen bij scotopisch omgevingslicht.[6]
De uitlopers van de UIT-bipolaire cellen vormen een synaps in de buitenste laag van de binnenste plexiforme laag van het netvlies en die van de AAN-bipolaire cellen eindigen in de binnenste laag van de binnenste plexiforme laag. De cellichamen van de retinale bipolaire cellen liggen ertussen in de binnenste granulaire laag.
Signaaloverdracht
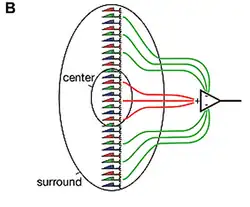
Retinale bipolaire cellen brengen effectief informatie over van staafjes en kegeltjes naar ganglioncellen. De horizontale cellen en de amacriene cellen compliceren de zaken enigszins. De horizontale cellen introduceren laterale inhibitie in de dendrieten en veroorzaken de centrumomgevings (surround) inhibitie die zichtbaar is in de receptieve velden van het netvlies. De amacriene cellen introduceren ook laterale inhibitie in het axonterminale gedeelte, dat verschillende visuele functies vervult, waaronder efficiënte signaaltransductie met een hoge signaal-ruisverhouding.[7]
Het mechanisme voor het creëren van het centrum van het receptieve veld van een retinale bipolaire cel is bekend: directe innervatie van de lichtgevoelige cel erboven, via een metabotrope (AAN) of ionotrope (UIT) receptor. Het mechanisme voor het creëren van de monochromatische omgeving (surround) van hetzelfde receptieve veld wordt echter onderzocht. Hoewel bekend is dat de horizontale cel een belangrijke cel in dit proces is, is de exacte volgorde van receptoren en moleculen onbekend.
De axonen van ganglioncellen vormen de binnenste laag van het netvlies als een laag zenuwvezels en, na het verlaten van het oog, de oogzenuw. Elk van deze ganglioncellen krijgt een specifiek gebied van het netvlies toegewezen als receptief veld door onderlinge verbinding via retinale bipolaire cellen.
AAN-centrumomgevingscellen verhogen hun activiteit wanneer licht op het centrum van hun receptieve veld valt, maar verlagen hun activiteit wanneer de omgeving van hun receptieve veld rondom het centrum wordt belicht. UIT-centrumomgevingscellen reageren precies andersom: wanneer licht op het centrum valt, verlagen ze hun activiteit; wanneer licht op de omgeving valt, verhogen ze hun activiteit.
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Retina bipolar cell op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.
- ↑ Tsukamoto Y and Omi N (2017) Classification of Mouse Retinal Bipolar Cells: Type-Specific Connectivity with Special Reference to Rod-Driven AII Amacrine Pathways. Front. Neuroanat. 11:92. doi: 10.3389/fnana.2017.00092
- ↑ Wenjun Yan, Yi-Rong Peng, Tavé van Zyl (2020). Cell Atlas of The Human Fovea and Peripheral Retina. Scientific Reports 10 (1): 9802. PMID 32555229. PMC 7299956. DOI: 10.1038/s41598-020-66092-9.
- ↑ Rikard Frederiksen, Yi-Rong Peng, Alapakkam P. Sampath (2025). Evolution of rod bipolar cells and rod vision. The Journal of Physiology. PMID 39775947. DOI: 10.1113/JP287652.
- ↑ T. Baden1,2 and D. Osorio1. The Retinal Basis of Vertebrate Color Vision. Vol. 5:177-200, June 21, 2019
- ↑ Principles of Cognitive Neuroscience. Sinauer Associates Inc, Sunderland, Mass (2007), p. 253. ISBN 978-0-87893-694-6.
- ↑ Bloomfield Stewart A., Dacheux Ramon F. (2001). Rod Vision: Pathways and Processing in the Mammalian Retina. Progress in Retinal and Eye Research 20 (3): 351–384. PMID 11286897. DOI: 10.1016/S1350-9462(00)00031-8.
- ↑ Tanaka M, Tachibana M (15 August 2013). Independent control of reciprocal and lateral inhibition at the axon terminal of retinal bipolar cells. J Physiol 591 (16): 3833–51. PMID 23690563. PMC 3764632. DOI: 10.1113/jphysiol.2013.253179.