Stereopsis (zien)
In de wetenschap van het zien is stereopsis de sensatie dat objecten in de ruimte niet plat zijn maar zich uitstrekken in de diepte, en dat objecten onderling op verschillende afstanden staan. Deze sensatie is veel sterker dan de suggestie van diepte die door tweedimensionaal perspectief wordt opgeroepen.
Het woord stereopsis komt van Griekse stereós, dat 'vast' betekent, en ópsis, dat 'uiterlijk, zicht' betekent. Samen duidt dit op het zien van de buitenkant van driedimensionale, "vaste" objecten.
Bij de mens zijn er twee mechanismen die de sensatie van stereopsis oproepen: binoculair dieptezien op basis van binoculaire dispariteit en monoculair dieptezien op basis van optische flow. Bij binoculair dieptezien ontstaat de sensatie door het verwerken van de verschillen in de retinabeelden die het gevolg zijn van het feit dat de twee ogen uit een andere richting kijken (binoculaire dispariteit). En bij bewegingszien ontstaat de sensatie door het verwerken van bewegingsinformatie als de waarnemer beweegt (optische stroom, parallax). De sensatie van stereopsis is in beide gevallen vergelijkbaar. Dit wordt in het onderstaande plaatje geïllustreerd. In het plaatje worden afwisselend de linker en rechter afbeelding van een stereofoto getoond. Mensen die op de foto dichterbij staan lijken sneller te bewegen dan mensen die verder weg staan. Dit wordt waargenomen als diepte: de personen lijken in diepte los van elkaar te staan. Als de twee afbeeldingen naast elkaar in een stereoscoop zouden worden bekeken dan wordt hetzelfde 3D beeld waargenomen, maar nu zonder beweging.
 Stereopsis veroorzaakt door alternerende stereobeelden.
Stereopsis veroorzaakt door alternerende stereobeelden.
In onderzoek naar dieptezien wordt de term stereopsis overwegend gebruikt voor binoculair dieptezien en niet voor de sensatie van diepte als gevolg van optische flow of parallax. Soms wordt de term relatieve diepte gebruikt. Deze term benadrukt dat niet de afstand tot de waarnemer wordt bedoeld maar de onderlinge diepterelaties van de waargenomen objecten. Als de betekenis duidelijk is uit de context, dan wordt ook het enkele woord diepte gebruikt in plaats van relatieve diepte.
Binoculair dieptezien komt voor in twee kwaliteiten: grove stereopsis en fijne stereopsis. Fijne stereopsis speelt een rol bij het herkennen van vormen en objecten en grove stereopsis bij ruimtelijke lokalisatie. In de hersenen zijn hiervoor twee neurofysiologische mechanismen aanwezig.
Binoculair dieptezien is een specialisatie van het vermogen tot richtingzien dat in een apart artikel wordt behandeld. Binoculair dieptezien is gebaseerd op kleine verschillen (dispariteiten) in de richting waarin het linker en rechter oog een object zien, en die het gevolg zijn van het feit dat de twee ogen ongeveer 6,5 cm uit elkaar staan.
Voorwaarden voor het optreden van binoculair dieptezien zijn dat de visuele richtingen in het linker en rechter oog een bepaalde gelijkenis hebben, min of meer op het zelfde moment worden geprikkeld, en het verschil tussen de richtingen in het linker en rechter oog (horizontale dispariteit) beperkt is. In het volgende wordt voor de genoemde aspecten op hoofdlijnen beschreven welke kennis er is over normale stereopsis bij de mens, en worden de basisbegrippen uitgelegd die nodig zijn om onderliggende brondocumenten te begrijpen..
Samenvatting onderzoek
Het onderzoek naar binoculair dieptezien begint bij Charles Wheatstone. Hij toonde aan het eind van de 19e eeuw als eerste aan dat horizontale dispariteit van verticale lijnen voldoende is om een sensatie van diepte op te roepen.[1][2][3][4] Bela Julesz toonde in de 20e eeuw aan dat de sensatie ook optreedt met stippen (random dot stereogram) en dat het zien van diepte vooraf gaat aan het waarnemen van vormen. Jodi Krol toonde rond dezelfde tijd aan dat een licht-overgang (rand) nodig is, maar dat twee corresponderende randen met tegengesteld contrast geen dieptesensatie geven. Hij vond ook dat de diepte van de oppervlakken tussen deze randen een interpretatie zijn.[5][6][7] De linker en rechter visuele richting moeten ongeveer gelijktijdig worden geprikkeld, maar dit hoeft niet exact op hetzelfde moment te gebeuren. Dit wordt geillustreerd met de Pulfrichillusie.
Kenneth Ogle (1950) vond dat de kwaliteit van de dieptesensatie verschilt bij kleine en bij grote dispariteiten en onderscheidt op basis hiervan verschillende typen stereopsis.[8][9] Er wordt algemeen aangenomen dat bij het ontbreken van een sensatie van stereopsis het waargenomen beeld meestal in het vlak van de horopter wordt gezien. John Foley (1972) beschrijft dat in uitzonderingsgevallen het beeld ook iets achter of voor de horopter kan verschijnen.[10] Jodi Krol (1982) laat zien dat dit laatste gebeurt als de ogen door bepaalde reflexen onbewust iets voor of achter het bedoelde fixatiepunt zijn gericht en het fixatiepunt dus niet op de horopter ligt.[7]
Ten slotte, in de visuele cortex zijn zenuwcellen gevonden die zijn afgestemd op een bepaalde dispariteit.[11] Deze cellen vormen de grondslag voor neurale modellen voor binoculair dieptezien[12][13] en voor de oplossing van het te bespreken correspondentieprobleem. Deze cellen zijn onderdeel van twee neurofysiologische mechanismen die zijn gespecialiseerd in respectievelijk het herkennen van vormen en objecten[14] en in ruimtelijke lokalisatie.[15]
Binoculaire dispariteit
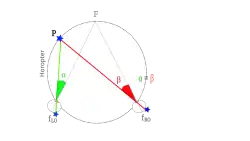
Als de aandacht wordt gericht op een punt F in de ruimte dan worden automatische oogbewegingen uitgevoerd waardoor het oog draait en punt F wordt afgebeeld op het punt van het oog waarmee het scherpst kan worden gezien, de fovea. De richting waarin het oog dan kijkt wordt de hoofdrichting van het oog (oculocentrische hoofdrichting) genoemd. Elk ander punt in de ruimte wordt door het oog in een bepaalde richting gezien die is uit te drukken als de hoek die deze richting maakt met de hoofdrichting. Dit wordt visuele richting of kortweg richting genoemd. De richtingen waarin elk oog hetzelfde object zien zijn soms gelijk, maar meestal niet. Het verschil in richtingen wordt dispariteit genoemd. In het aparte artikel over richtingzien wordt uitgelegd hoe de hersenen de richtingen die elk oog ziet combineren tot een gecombineerd beeld met enkelbeelden, dubbelbeelden en gefuseerde beelden die schijnbaar worden gezien vanuit een punt midden tussen beide ogen (cyclopisch oog).
Horizontale dispariteit
De horizontale afstand van ca 6,5 cm tussen de twee ogen zorgt er voor dat punten in de ruimte die op verschillende dieptes ten opzichte van het fixatiepunt liggen, een horizontaal richtingsverschil hebben. Dit verschil wordt horizontale dispariteit genoemd en uitgedrukt als het verschil tussen de hoeken α en β in de figuur. Naast horizontale dispariteit bestaat er ook verticale dispariteit. Met deze term wordt aangegeven dat de ogen verticaal niet zijn uitgelijnd bijvoorbeeld doordat het hoofd niet precies verticaal staat. Dit laatste wordt meestal gedeeltelijk of geheel gecorrigeerd met automatisch uitgevoerde oogbewegingen . Verticale dispariteiten kunnen in sommige gevallen ook een dieptegevoel oproepen.[16]
De termen horizontale en verticale dispariteit zijn alleen logisch als de waarnemer zich rechtop bevindt. Een andere term die soms wordt gebruikt en die de lading beter dekt is binoculaire dispariteit. Binoculaire dispariteit heeft veel overeenkomsten met bewegingsparallax en wordt daarom soms ook wel binoculaire parallax genoemd. Ook bij beweginsparallax wordt een dieptesensatie opgeroepen, maar deze vereist dat de waarnemer beweegt of de waargenomen objecten bewegen ten opzichte van elkaar. Onder neurofysiologische mechanismen is te lezen dat bewegingsparallax mogelijk in het zelfde systeem wordt verwerkt als grove stereopsis.
Horopter
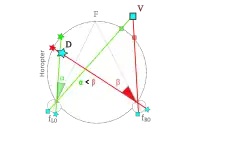
Punten op gelijke diepte met het fixatiepunt liggen op een cirkel door beide ogen en het fixatiepunt. Deze cirkel wordt horopter genoemd. Punten op deze cirkel projecteren op corresponderende punten in beide ogen, d.w.z. op punten die in het linker en rechter oog in dezelfde richting kijken. De horopter nemen we waar als een soort van scherm waarop we de wereld zien.
Gekruiste dispariteit
De richtingen in beide ogen van een punt D dat dichter bij is dan de horopter worden op dit scherm gekruist gezien: de richting waarin de richting van het linker oog wordt gezien bevindt zicht rechts van de richting van het rechter oog. Dit wordt gekruiste dispariteit of negatieve dispariteit genoemd.
Parallele dispariteit
De richtingen in beide ogen van een punt V dat verder weg is dan de horopter worden op dit scherm naast elkaar gezien: de rechter richting wordt rechts gezien. Dit wordt parallelle dispariteit of positieve dispariteit genoemd.
Bewijs van Wheatstone
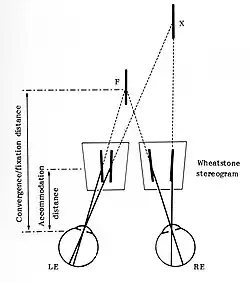
Charles Wheatstone toonde in 1838 aan dat twee objecten die (alleen) verschillen in horizontale dispariteit, een sensatie van relatieve diepte oproepen.[17] Om te zorgen dat deze sensatie niet door tweedimensionaal perspectief kon zijn veroorzaakt gebruikte hij twee platte plaatjes met elk twee vertikale lijnen, een lijnstereogram, om de visuele richtingen op te roepen en vond hij de stereoscoop uit om deze plaatjes te kunnen bekijken.
Diepte als functie van dispariteit
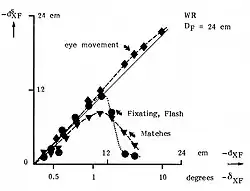
Ogle (1950) heeft onderzocht hoe de sensatie van diepte af hangt van mate van dispariteit.[18] Ogle vroeg een waarnemer om een punt F te fixeren en bewoog dan een verticale staaf Y in diepte weg van X. De grootte van de waargenomen diepte bleek bij kleine dispariteiten eerst lineair toe te nemen, en bij grotere dispariteiten weer af te nemen, en bij nog grotere dispariteiten totaal afwezig te zijn. In het laatste geval werd Y op de afstand van de horopter gezien.
Als Ogle de waarnemer vroeg tussen X en Y heen en weer te kijken (vergeren), dan bleek in een veel groter gebied dieptezien mogelijk.

Volgens het onderzoek van Ogle en anderen worden diepteverschillen alleen waargenomen in een smal gebied in de ruimte, verder weg en dichterbij dan de horopter en gebeurt dit met drie kwaliteiten: fijne stereopsis, grove stereopsis en geen stereopsis.[19][20]
Fijne stereopsis
Bij fijne stereopsis kan de waarnemer de grootte van de diepte aangeven: hoe groter de dispariteit, hoe groter de waargenomen diepte. Ogle gebruikte hiervoor de term patente stereopsis. Latere onderzoekers vervingen deze term door fijne stereopsis.
Grove stereopsis
Bij grove stereopsis kan de waarnemer wel aangegeven of het ene object verder weg staat dan het andere, maar niet hoeveel. Ogle gebruike hiervoor de term kwalitatieve stereopsis. Latere onderzoekers vervingen deze term door grove stereopsis.
Geen stereopsis
In het overblijvende gebied worden objecten zonder dieptesensatie gezien, op of rond de afstand van de horopter, en meestal als dubbelbeeld.
Voorwaarden voor binoculaire diepte
Horizontale dispariteit is een voorwaarde voor het zien van relatieve diepte, maar niet een voldoende voorwaarde. De visuele richtingen in het linker en rechter oog moeten ook een bepaalde gelijkenis hebben zoals gelijke licht-overgangen (randen) of vlekjes, en moeten min of meer gelijktijdig worden geprikkeld. Oogbewegingen zorgen er daarbij voor dat objecten in de ruimte zo veel mogelijk op corresponderende punten in beide ogen vallen en de dispariteit zo klein mogelijk blijft.
Gelijke randen
Hering suggereerde al in 1864 dat de visuele richtingen gelijke "randen" moeten bevatten en dat de oppervlakken tussen deze randen bij het waarnemen worden "ingevuld".[21] Dit is door Krol (1982, p. 38-39) bevestigd.[22] Randen met een tegengesteld contrast geven geen dieptesensatie. Het zien van een tegengesteld contrast wekt een oogbewegingsreflex op waardoor de tegengestelde contrasten niet op corresponderende punten vallen en de randen iets voor of achter het fixatiepunt lijken te liggen.[7]
Kleur en helderheid
De kleur en helderheid hoeft niet gelijk te zijn. Stereopsis treedt zelfs op bij tegengestelde kleuren, waarbij de waargenomen kleur afwisselt tussen de twee kleuren. Er is daarbij geen kleurmenging (Krol 1982, p. 38-39).[22]
Random dot stereogram
Julesz (1971) heeft met random dot stereogrammen aangetoond dat stereopsis ook plaats vindt als de visuele richtingen bestaan uit kleine, willekeurige witte en zwarte vlekjes, waarbij een bepaalde groep vlekjes in het linker beeld is verschoven in het rechter oog. Ieder oog afzonderlijk ziet alleen maar vlekjes, maar beide ogen samen zien de groep van verschoven vlekjes als een vorm met diepte.[23]


Figuur. Random dot stereogram

Figuur. Verborgen vorm. In het beeld worden afwisselend het linker en rechter plaatje van het bovenstaande random dot stereogram getoond. Door de beweging die dan zichtbaar wordt is te zien welke vorm in het stereogram verborgen is. In dit geval een vierkant. Dit vierkant wordt ook zichtbaar als het stereogram met een stereoscoop wordt bekeken, alleen dan beweegt het vierkant niet.
Gelijktijdigheid
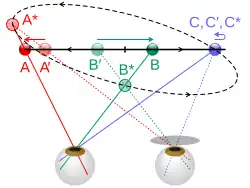
De visuele richting in het linker en rechter oog moeten gelijktijdig of kort na elkaar geprikkeld worden. Een illustratie van niet-gelijktijdige prikkeling is de Pulfrich illusie, waarbij (volgens de verklaring van het Pulfrich effect) een donker glas voor een van beide ogen de signalen van dat oog vertraagt.
Bij het bekijken van de heen en weer gaande beweging van de ruitenwisser van een auto, met een grijsfilter of een zonnebrilglas voor een van de ogen, lijkt de ruitenwisser een elliptische beweging in diepte te maken. Hij lijkt zelfs door het glas heen te bewegen. Deze illusie werd voor het eerst beschreven door Pulfrich in 1922.
Oogbeweging
Oogbewegingen zijn belangrijk om de voorwaarden die nodig zijn voor stereopsis te realiseren en deze voorwaarden in stand te houden. Verticale oogbewegingen en rotatiebewegingen zorgen er voor dat bij een beweging van het hoofd de beelden in het linker en rechter oog op de juiste, gelijke hoogte komen en blijven. Vergentiebewegingen (convergentie en divergentie) zorgen er voor dat de horopter zodanig in de ruimte wordt geplaatst en gehouden dat objecten die de aandacht hebben, optimaal binnen het gebied voor stereopsis liggen.[24] Een groot deel van deze bewegingen gebeurt reflexmatig.
Controle op juiste fixatie
Om bij onderzoek te zorgen dat de ogen het juiste punt fixeren (vergentie) en reflexen het resultaat niet beïnvloeden, worden verschillende methoden toegepast. Een methode bestaat uit het onverwacht en dusdanig kort aanbieden van de stimuli dat de ogen geen tijd hebben om een oogbewegingsreflex uit te voeren. Een andere methode bestaat uit het met zogenaamde nonius lijnen zichtbaar maken dat de waarnemer juist fixeert. Het linker en rechter oog zien daarbij, boven respectievelijk onder het fixatiepunt, allebei een haarlijn die het andere oog niet ziet en die uitgelijnd moeten zijn. Een laatste methode bestaat uit het registreren van de oogbewegingen tijdens het experiment.
Correspondentieprobleem
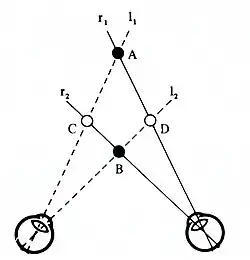
Als er in een oog een visuele richting is die met meerdere visuele richtingen in het andere oog gecombineerd kan worden om een sensatie van diepte te geven, dan staan de hersenen voor een keuzeprobleem: welke combinatie is de juiste en wordt er gezien? Julesz noemt dit het correspondentieprobleem en heeft dit probleem met Random dot-stereogrammen onderzocht.[23] Uit dit onderzoek blijkt o.a. dat het visuele systeem de voorkeur geeft aan het zien van vlakken (globaliteitsprincipe) en dat vormen pas worden ingevuld nadat dispariteiten zijn vastgesteld. Krol heeft het correspondentieprobleem ook onderzocht, maar nu met natuurlijke stimuli (tweespijkerillusie). waarbij door te voelen en uit meerdere posities te kijken, kan worden vastgesteld welke "interpretatie" de juiste is. Het blijkt dat altijd de interpretatie met de minste diepte wordt gezien.
Binoculaire schijnbeelden
Bij de tweespijkerillusie worden twee naalden op de posities A en B getoond. Onder de juiste waarnemingscondities zijn deze naalden onzichtbaar en worden in plaats van deze echte objecten twee naalden op de posities C en D gezien. Deze naalden zijn er niet echt en worden door Krol daarom schijnbeelden genoemd.
Neurofysiologische mechanismen
In de jaren zestig ontdekten Horace Barlow, Colin Blakemore en Jack Pettigrew neuronen in de visuele cortex van de kat waarvan het receptieve veld zich in verschillende horizontale posities in beide ogen bevond.[25] Hiermee werd de neurale basis voor stereopsis gelegd. Hun bevindingen werden betwist door David Hubel en Torsten Wiesel, hoewel ze uiteindelijk toegaven toen ze vergelijkbare neuronen vonden in de visuele cortex van de aap.[26] In de jaren 80 ontdekten Gian Poggio en anderen neuronen in de V2 van de apenhersenen die reageerden op de diepte van stereogrammen met willekeurige stippen.[27]
Fijne stereopsis wordt verwerkt door een systeem van kleine cellen dat fijne details en kleur verwerkt in een omgeving die relatief langzaam verandert.[14] Dit systeem speelt een grote rol in het herkennen van vormen en objecten en houdt zich bezig met "wat" wordt gezien. Grove stereopsis (en daaraan gekoppelde dubbelbeelden) worden verwerkt door een systeem van grote cellen dat grote veranderingen zoals beweging, contrast en snelle veranderingen verwerkt in een relatief snel veranderende omgeving.[15] Dit systeem is gericht op localisatie en oriëntatie in de ruimte. De zenuwbanen van beide systemen lopen van de ogen naar de visuele cortex. Het systeem voor fijne stereopsis loopt vervolgens door naar de inferieure temporale cortex waar "wat"-informatie wordt verwerkt (parvo route). Het systeem voor grove steropsis loop door naar de posterieure pariëtale cortex waar de “waar”- en “hoe”-informatie wordt verwerkt (magno route). Beide systemen lijken gedeeltelijk parallel aan elkaar te werken maar werken ook samen. Als een systeem uitvalt kan het andere systeem soms nog wel functioneren: mensen die geen fijne stereopsis hebben kunnen soms wel grove stereopsis ervaren.[28] Voor dynamische dispariteitsverwerking, zie[29][30][31]
Voordelen
Stereopsis heeft een positief effect op het oefenen van praktische taken zoals het inrijgen van een naald en het vangen van ballen (met name bij snelle balspellen).[32]), het uitschenken van vloeistoffen en andere. Beroepsactiviteiten kunnen het bedienen van stereoscopische instrumenten zoals een binoculaire microscoop omvatten. Hoewel sommige van deze taken kunnen profiteren van compensatie van het visuele systeem door middel van andere dieptesignalen, zijn er enkele functies waarvoor stereopsis noodzakelijk is. Beroepen die een nauwkeurige afstandsbeoordeling vereisen, vereisen soms een zekere mate van stereopsis; vliegtuigpiloten hebben in het bijzonder zo'n vereiste (zelfs als de eerste piloot die solo rond de wereld vloog zijn prestatie volbracht met alleen monoculair zicht.)[33] Ook chirurgen[34] vertonen normaal gesproken een hoge stereoacuïteit. Wat betreft autorijden vond één onderzoek een positief effect van stereopsis in specifieke situaties, alleen op middellange afstanden.[35] Bovendien vond een onderzoek onder ouderen dat schittering, verlies van gezichtsveld en het bruikbare gezichtsveld voorspellers waren van betrokkenheid bij een ongeval, terwijl de visuele scherpte, contrastgevoeligheid en stereoacuïteitsscores van ouderen niet geassocieerd waren met ongevallen.[36]
Binoculair zien heeft naast stereopsis nog andere voordelen, met name de verbetering van de kwaliteit van het zicht door binoculaire sommatie; Mensen met scheelzien (zelfs degenen zonder diplopie) scoren lager op binoculaire sommatie, en dit lijkt mensen met scheelzien ertoe aan te zetten om in visueel veeleisende situaties één oog te sluiten.[37][38] Stereopsis is ook belangrijk voor objectherkenning en voor het doorzien van camouflage.[23]
Bij dieren
Zien is een vorm van natuurlijke intelligentie[39] die mogelijkheden biedt om te oriënteren in de ruimte, om objecten te herkennen, objecten te manipuleren en om te navigeren. Of en hoe deze mogelijkheden zich binnen de mens en andere diersoorten hebben ontwikkeld, is een vraag voor de biologie en de ecologische psychologie. James Gibson beschrijft in de ecologie van het zien dat deze mogelijkheden zijn ontwikkeld in een interactief samenspel tussen waarnemer en zijn omgeving en dat de omgeving vaak veel meer praktisch bruikbare informatie bevat dan we denken.[40] De beschikbare informatie kan per dier en dus leefomgeving verschillen. Bijvoorbeeld, piloten maar ook vogels die zich snel verplaatsen hebben genoeg aan (monoculair) bewegingszien om te kunnen sturen en objecten te kunnen vermijden. Bij de mens leidt dit zelfs tot een monoculaire sensatie van stereopsis. Binoculair dieptezien voegt hier aan toe dat deze sensatie ook beschikbaar is in situaties waarin de waarnemer niet heel snel beweegt. Dit kan belangrijk zijn voor dieren die vruchtjes in een struik willen pakken, die stilletjes op een naderende prooi wachten totdat deze binnen grijpafstand is, of die zich zich snel zonder struikelen over een hobbelig terrein willen bewegen.
Of dieren met overlappende gezichtsvelden een vorm van binoculair dieptezien hebben kan in theorie op verschillende manieren worden vastgesteld. Zo kan worden nagegaan of dieren binoculaire dispariteit gebruiken om te kunnen bepalen of een object dichtbij genoeg is om het te kunnen "grijpen" of te manipuleren. Ook kunnen hersengolven worden gemeten en worden nagegaan of hierin de reactie op een random dot stereogram hetzelfde is als bij mensen. En bij het uitvoeren van natuurlijke taken kan worden nagegaan of de taakuitvoering wordt vertraagd en meer fouten geeft als dispariteitinformatie wordt veranderd.
Bij onderzoek naar de aanwezigheid van binoculair zien zou onderscheid moeten worden gemaakt tussen het zien van absolute of egocentrische diepte, binoculaire relatieve diepte (stereopsis) en monoculaire relatieve diepte (parallax). Bij de demonstratie van stereopsis wordt dit onderscheid niet altijd gemaakt; het is in dat geval niet mogelijk om te concluderen welke vorm van dieptezien aanwezig is.
Egocentrische diepte

Padden en bidsprinkhanen lijken de absolute afstand tot hun prooi (egocentrische diepte) te kunnen zien: als met prisma's de schijnbare afstand waarop ze hun prooi zien wordt veranderd, dan "grijpen" ze mis.[41] Dit wordt verklaard door aan te nemen dat deze dieren via een soort van driehoeksmeting afstand kunnen bepalen. Driehoeksmeting is in theorie mogelijk doordat de ogen van padden en bidsprinkhanen een vaste positie in het hoofd hebben en niet kunnen bewegen, waardoor de hoeken waaronder het linker en rechter oog een object zien samen de afstand tot het object bepalen. Dit is echter gecompliceerder dan het lijkt. De via driehoeksmeting bepaalde egocentrische afstand is namelijk afhankelijk van de afstand tussen de beide ogen (IOA). Hoe kent het organisme deze afstand? Deze afstand verandert bovendien gedurende de groeiperiode, wanneer ook de kop groter wordt. Hoe weet het organisme hoeveel de kop is gegroeid? Het organisme kan dit niet van nature weten, maar kan dit alleen leren op basis van gelukte en mislukte "grijp"-bewegingen. Een andere vraag is of padden en bidsprinkhanen naast de grijpafstand ook andere afstanden kunnen zien. Misschien hebben ze in de hersenen alleen een "vliegdetector-neuron" welke actief wordt als een vlieg zich binnen bereik bevindt, waarna een motorische reflex zorgt dat deze gegrepen wordt. Anders gezegd, misschien hebben ze dus geen bewuste dieptewaarneming. Dit is omdat een dier niet kan praten moeilijk vast te stellen.
De mens en sommige dieren hebben beweegbare ogen. Dit maakt ket kunnen detecteren van egocentrische diepte complexer dan bij dieren met vaste ogen. In principe zou de oogstand kunnen worden afgeleid uit informatie van de receptoren in de oogspieren (spierspoeltjes). Mensen blijken deze informatie niet te gebruiken om egocentrische diepte te zien, mensen zijn daar niet goed in.[42] Toch lijkt het alsof mensen wel goed egocentrische diepte kunnen zien. Immers een mens is in staat iets wat hij ziet direct op de juiste afstand met een vinger aan te raken. Dit komt dan omdat de relatieve diepte tussen het doelobject en de vinger gebruikt wordt om de hand te sturen.
Relatieve diepte
Stereopsis is een bepaalde vorm van dieptezien waarbij het ene punt dichterbij lijkt dan het andere (relatieve diepte) zonder dat het op een bepaalde afstand van de waarnemer wordt gezien, net als bij bewegingsparallax. Het verschil met bewegingsparallax is dat de relatieve diepte ook kan worden gezien als er geen of weinig beweging aanwezig is, dus als de waarnemer en de omgeving niet ten opzichte van elkaar bewegen. Vaak wordt benadrukt dat stereopsis belangrijk is voor de jagers in het dierenrijk omdat prooidieren zich verbergen door niet te bewegen en het hebben van camouflage. Echter er zijn ook andere evolutionaire voordelen zoals het lopen of rennen door hobbelig terrein zonder te struikelen. Dit zou het bestaan van stereopsis bij paarden en schapen kunnen verklaren. Of het kunnen grijpen van een prooi tijdens de vlucht door een roofvogel, maar ook het gebruik van kunstmatige instrumenten. Zo blijkt een kraai die als een van de weinige vogels een stokje als instrument gebruikt, over een vorm van stereozien te beschikken in het gebied in de ruimte waar het stokje wordt gebruikt.[43]
De plaats en de omvang van het gebied in de ruimte waarin diepte wordt gezien verschilt van dier tot dier en lijkt te zijn aangepast aan de ecologische omgeving. Ook over de nauwkeurigheid waarmee diepte wordt gezien is, behalve bij de mens weinig bekend. Laat staan over of en hoe diepte wordt waargenomen en welke mechanismen hieraan ten grondslag liggen. Alleen bij bepaalde apen en katten is aangetoond dat deze bij het zien van een random dot stereogram een zelfde soort hersengoven aanwezig is als bij de mens, hetgeen zou kunnen betekenen dat eenzelfde soort van neuronaal systeem actief is.
Demonstratie van stereopsis
Mede doordat er vele monoculaire dieptekenmerken zijn en dieren niet kunnen vertellen wat ze waarnemen, is het bijzonder moeilijk om eenduidig aan te tonen dat dieren dieptezien hebben dat lijkt op het dieptezien bij de mens (stereopsis). Met uitzondering van bepaalde apen, katten en uilen is er weinig bekend over de capaciteit, functie en neuronale basis voor dieptezien bij dieren.[41]
Er zijn aanwijzingen voor de aanwezigheid van een vorm van binoculair dieptezien[44] aangetroffen bij veel gewervelde dieren[44] , waaronder zoogdieren zoals paarden,[45] vogels zoals valken[46] en uilen,[47] reptielen, amphibia inclusief padden[48] en vissen. Het is ook aangetroffen bij ongewervelden[44] waaronder koppotigen zoals de inktvis,[49] schaaldieren, spinnen en insecten zoals bidsprinkhaan.[50] Stomatopoden hebben bijzonder ontwikkelde ogen. Ze kunnen een oog verliezen en hebben dan nog steeds stereopsis met slechts één oog.[51][44]
Samenvattend, vormen van binoculair dieptezien zijn gevonden in zoogdieren, vogels, amfibieën en insecten.[41] Binoculair zien is niet alleen van belang voor het doorzien van camouflage, maar ook voor het sturen van fysieke bewegingen. De plaats in de ruimte waar dieren diepte kunnen zien verschilt per dier en is afhankelijk van het natuurlijke gedrag. Ook de grootte van het gebied verschilt, waarbij dieren met de ogen van voren waarschijnlijk in een groter gebied diepte kunnen zien dan dieren die de ogen van opzij hebben. De mens ziet (bijna) alleen relatieve diepte. Hoe dit is bij dieren is moeilijk vast te stellen.
Zie ook
- ↑ Wheatstone, C. (1838), "On some remarkable and hitherto unobserved phenomena of binocular vision", Philos. Trans. R. Soc. London Part 1,371 – 395.
- ↑ Hering, E. (1864),"Vom binocularen tiefensehen"), In: Beiträge zur Physiologie. Fünftes heft. Wilhelm Engelmann: Leipzig. Erster Band. Leipzig. Thieme, 1931.
- ↑ Panum, PL. (1858), "Physiologische Untersuchingen über das Sehen mit zwei Augen" Kiel.
- ↑ Fröhlich (1895) "Unter welchem umstände erscheinen doppelbilder in ungleichen Abständen vom Beobachter?" Albrecht vom Graefes Arch. Klin. Exp. Ophtalm. 41, 134 157.
- ↑ Krol, JD, van de Grind, WA (1979), "On the sufficiency of disparate contours as a stimulus for stereopsis", Neurosc. Lett. Suppl. 3 S 304.
- ↑ Krol, JD, van de Grind, WA (1981),"Fixation disparities and depth vision due to a process of vergence fine-tuning", In: Proc. 22nd meeting of the Dutch Federation of Medical Scientific Societies, Utrecht, April 24, No. 248.
- 1 2 3 Krol, JD, van de Grind, WA (1982), ”Depth from dichoptic edges depends on vergence fine-tuning.”
- ↑ Ogle KN (1953), "Precision and validity of stereoscopic depth perception from double images", J. Opt. Soc. Am. 43, 906 – 513.
- ↑ Blakemore, C (1970) "The range and scope of binocular depth discrimination in man", J. of Pfysiol. Lond. 211, 599-622.
- ↑ Foley, JM, Richards, W (1972), "Effects of voluntary eye movement and convergence on the binocular appreciation of depth", Perception and Psychophysics 11, 423 – 427.
- ↑ Bishop, PO (1973), "Neurophysiology of binocular single vision and stereopsis", In: Handbook of sensory physiology, vol. VII/3A: Central processing of visual information. A: Integrative functions and comparative data.
- ↑ Sperling, G (1970), "Binocular vision: a physical and neural theory", Am. J. of Psychol. 83, 461 – 533.
- ↑ Krol, JD, van de Grind, WA (1989), "The double-nail illusion, experiments on binocular vision with nails needles and pins", Perception. 1980; 9(6): 651-69,
- 1 2 Wilcox LM, Allison RS (november 2009). Grof-fijn dichotomieën in menselijke stereopsis. Vision Research 49 (22): 2653–65. PMID 19520102. DOI: 10.1016/j.visres.2009.06.004.
- 1 2 Stidwill, David, Fletcher, Robert (8 november 2010). Normaal binoculair zicht: theorie, onderzoek en praktische aspecten. John Wiley & Sons, p. 164. ISBN 978-1-4051-9250-7.
- ↑ Matthews N;Meng X.; Xu P; Qian Q. (2003) “A physiological theory of depth perception from vertical disparity”, Vision Research. Volume 43, Issue 1, January 2003, Pages 85-99.
- ↑ Bijdragen aan de fysiologie van het zien. – Deel 1. Over enkele opmerkelijke en tot nu toe onopgemerkte verschijnselen van binoculair zien. Door CHARLES WHEATSTONE, F.R.S., hoogleraar experimentele filosofie aan King's College, Londen.
- ↑ Ogle K.N. (1950), "Researches in binocular vision", Philadelphia: Saunders.
- ↑ Ogle K.N. (1952a) "On the limits of stereoscopic vision" K, J. Exp. Psychol. 44, 253 -259.
- ↑ Ogle K.N. (1952b) "Disparity limits of stereopsis", A.M.A. Ophtalm. 48, 50- 60.
- ↑ Vom binocularen tiefensehen. E. Hering (1864), In: Beiträge zur Physiologie. Fünftes heft. Wilhelm Engelmann: Leipzig. Erster Band. Leipzig. Thieme, 1931.
- 1 2 Krol J.D. (1982), "Perceptual ghosts in stereopsis, a ghosly problem in binocular vision", PhD thesis ISBN 90-9000382-7.
- 1 2 3 Julesz B (1971),"Foundations of cyclopic perception", Chicago: University of Chicago Press,Foundations of cyclopean perception
- ↑ Harrold A.L.; Grove P.M. (2021), "The vergence horopter", Vision Research Volume 180, March 2021, Pages 63-79 DOI:10.1016/j.visres.2020.12.003
- ↑ Barlow HB, Blakemore C, Pettigrew JD (november 1967). Het neurale mechanisme van binoculaire dieptediscriminatie. The Journal of Physiology 193 (2): 327–42. PMID 6065881. PMC 1365600. DOI: 10.1113/jphysiol.1967.sp008360.
- ↑ Hubel DH, Wiesel TN (January 1970). Stereoscopisch zicht bij makaak. Cellen die gevoelig zijn voor binoculaire diepte in gebied 18 van de cortex van makaak. Nature 225 (5227): 41–2. PMID 4983026. DOI: 10.1038/225041a0.
- ↑ Poggio GF, Motter BC, Squatrito S, Trotter Y (1985). Reacties van neuronen in de visuele cortex (V1 en V2) van de alerte makaak op dynamische stereogrammen met willekeurige stippen. Vision Research 25 (3): 397–406. PMID 4024459. DOI: 10.1016/0042-6989(85)90065-3.
- ↑ Zie de interpretatie van uitspraken van Bela Julesz in: Leonard J. Press: The Dual Nature of Stereopsis – Deel 6 (gedownload op 8 september 2014)
- ↑ (2009). Onopgeloste problemen in stereopsis: dynamische dispariteitsverwerking. Spatial Vision 22: 83–90. PMID 19055888. DOI: 10.1163/156856809786618510.
- ↑ Ban H, Preston TJ, Meeson A, Welchman AE (februari 2012). De integratie van bewegings- en dispariteitssignalen met diepte in de dorsale visuele cortex. Nature Neuroscience 15 (4): 636–43. PMID 22327475. PMC 3378632. DOI: 10.1038/nn.3046.
- ↑ (augustus 1999). Het modelleren van de combinatie van bewegings-, stereo- en convergentiehoeksignalen met visuele diepte. Neural Computation 11 (6): 1297–330. PMID 10423497. DOI: 10.1162/089976699300016250.
- ↑ Mazyn LI, Lenoir M, Montagne G, Savelsbergh GJ (Augustus 2004). The contribution of stereo vision to one-handed catching. Experimental Brain Research 157 (3): 383–90. PMID 15221161. DOI: 10.1007/s00221-004-1926-x.
- ↑ Elshatory YM, Siatkowski RM (2014). Wiley Post, Around the World Without Stereopsis. Survey of Ophthalmology 59 (3): 365–72. PMID 24359807. DOI: 10.1016/j.survophthal.2013.08.001.
- ↑ Biddle M, Hamid S, Ali N (February 2014). An Evaluation of Stereoacuity (3D Vision) in Practicing Surgeons in various surgical specialties. De Chirurg 12 (1): 7–10. PMID 23764432. DOI: 10.1016/j.surge.2013.05.002.
- ↑ Bauer A, Dietz K, Kolling G, Hart W, Schiefer U (July 2001). De relevantie van stereopsis voor bestuurders: een pilotstudie. Graefe's Archive for Clinical and Experimental Ophthalmology 239 (6): 400–6. PMID 11561786. DOI: 10.1007/s004170100273.
- ↑ Rubin GS, Ng ES, Bandeen-Roche K, Keyl PM, Freeman EE, West SK (april 2007). Een prospectief, populatiegebaseerd onderzoek naar de rol van visuele beperkingen bij verkeersongevallen onder oudere bestuurders: de SEE-studie. Investigative Ophthalmology & Visual Science 48 (4): 1483–91. PMID 17389475. DOI: 10.1167/iovs.06-0474.
- ↑ Pineles SL, Velez FG, Isenberg SJ, Fenoglio Z, Birch E, Nusinowitz S, Demer JL (November 2013). Functionele werklast van scheelzien: verminderde binoculaire sommatie en binoculaire inhibitie. JAMA Ophthalmology 131 (11): 1413–9. PMID 24052160. PMC 4136417. DOI: 10.1001/jamaophthalmol.2013.4484.
- ↑ Damian McNamara, "Studie naar scheelzien onthult visuele functietekorten", 23-09-2013.
- ↑ Grind WA van de (2007),"Natuurlijke intelligentie", ISBN 9789057121869
- ↑ Gibson, J.J. (1950). De perceptie van de visuele wereld. Houghton Mifflin.
- 1 2 3 Nityananda V, Read JCA (2017), “Stereopsis in animals: evolution, function and mechanisms”, J Exp Biol, 2017 Jul 15;220(14):2502–2512. doi: 10.1242/jeb.143883
- ↑ Bradshaw MF, Parton AD, Glennerster A (2000), "The task-dependent use of binocular disparity and motion parallax information", Vision Res. 40, 3725-3734. 10.1016/S0042-6989(00)00214-5 [DOI] [PubMed] [Google Scholar]
- ↑ Martin G.R. (2017), “The sensory ecology of birds.”, Oxford University Press, ISBN 978-0-19-969453-2, doi 10.1093/acprof:oso/9780199694532.001.00001
- 1 2 3 4 Howard IP, Rogers B (1995),"Binocular vision and stereopsis",Oxford University Press, New York
- ↑ Timney, Brian, Keil, Kathy (1999). Lokale en globale stereopsis bij het paard. Vision Research 39 (10): 1861–1867. DOI: 10.1016/S0042-6989(98)00276-4.
- ↑ Fox, Robert, Lehmkuhle, Stephen W., Bush, Robert C. (1977). Stereopsis in de Falcon. Science 197 (4298): 79–81. ISSN: 0036-8075. DOI: 10.1126/science.867054.
- ↑ van der Willigen, R. F. (10 juni 2011). Uilen zien in stereo net zoals mensen. Journal of Vision 11 (7): 10–10. ISSN: 1534-7362. DOI: 10.1167/11.7.10.
- ↑ Collett, T. (1977). Stereopsis bij padden. Nature 267 (5609): 349–351. ISSN: 0028-0836. DOI: 10.1038/267349a0.
- ↑ Feord, R. C., Sumner, M. E., Pusdekar, S., Kalra, L., Gonzalez-Bellido, P. T. (10 januari 2020). Inktvissen gebruiken stereopsis om prooien aan te vallen. Science Advances 6 (2). ISSN: 2375-2548. PMID 31934631. PMC 6949036. DOI: 10.1126/sciadv.aay6036.
- ↑ Rossel, Samuel (1983). Binoculaire stereopsis bij een insect. Nature 302 (5911): 821–822. ISSN: 0028-0836. DOI: 10.1038/302821a0.
- ↑ Fox, Helen, Hoe beter je kunt zien met.... University of California Museum of Paleontology (2001). Geraadpleegd op 2 maart 2021.