Manipulonyx

Manipulonyx is een geslacht van theropode dinosauriërs, behorende tot de Maniraptora, dat tijdens het late Krijt leefde in het gebied van het huidige Mongolië. De enige benoemde soort is Manipulonyx reshetovi.
Vondst en naamgeving
In 1979 vond de Russische paleontoloog Walerij Joerjewitsj Resjetow (Валерий Юрьевич Решетов) bij de Chermeen Tsaw een gedeeltelijk skelet van een alvarezsauride. Het werd toegevoegd aan de collectie van het paleontologisch instituut van Moskou.
In 2025 werd de typesoort Manipulonyx reshetovi benoemd en beschreven door Aleksandr Olegowitsj Awerianow, Alksej Wladimirowitsj Lopatin en Andrej Anatoliwitsj Atoetsjin. De geslachtsnaam combineert het Latijn manipulare, "hanteren" met het Grieks onyx, "klauw", een verwijzing naar de hypothese van de beschrijvers dat de bijzondere bouw van de hand van de alvarezsauriden een aanpassing is aan het manipuleren van geroofde eieren. De soortaanduiding eert Resjetow als ontdekker. De Life Science Identifiers, de registraties bij de zoobank, zijn 6205BAAC-73384E73-A40D-D56BF5586638 voor het geslacht en 7F0428F3-BCF6-4D9A-A7AA-EEF9E36CABC6 voor de soort.
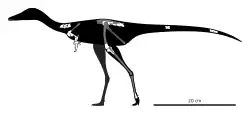
Het holotype, PIN 3142/364, is gevonden in een laag van de Nemegtformatie die dateert uit het Maastrichtien. Het bestaat uit een gedeeltelijk skelet zonder schedel. Bewaard zijn gebleven twee achterste halswervels, de voorste ruggenwervel, vier sacrale wervels, talrijke staartwervels, de linkerschoudergordel, beide opperarmbeenderen, de complete rechteronderarm inclusief pols en hand die zo meer volledig bekend is dan van enig andere directe verwant, het rechterdarmbeen, het rechterschaambeen en stukken uit alle delen van beide achterpoten.
Beschrijving
Grootte en onderscheidende kenmerken
Het holotype is een klein dier, zo'n een tot anderhalve meter lang.
De beschrijvers stelden een unieke combinatie van op zich niet unieke kenmerken vast. Het opperarmbeen is kort en robuust. De binnenhoek van de bovenkant van het opperarmbeen steekt boven de humeruskop uit. Er bevindt zich geen inkeping tussen de humeruskop en de deltopectorale kam. De deltopectorale kam van het opperarmbeen is vergroot. Het opperarmbeen heeft maar één enkele gewrichtsknobbel voor het contact met de onderarm, dus met de ellepijp en het spaakbeen samen. De buitenste beenstijl van het opperarmbeen richting de gewrichtsknobbel is sterk vergroot en steekt voorbij het gewricht uit. De processus olecrani van de ellepijp is sterk vergroot. De eerste vinger is sterk vergroot met een zeer grote en robuuste duimklauw. Het uitsteeksel op de bovenste voorkant van het eerste kootje van de eerste vinger is groot. De kanalen in de achterzijde van het tweede kootje van de eerste vinger zijn gesloten aan de buitenzijde. De tweede en derde vinger zijn sterk gereduceerd. De crista supraacetabularis, een horizontale richel op het darmbeen, is gewelfd en steekt niet verder naar voren uit dan het aanhangsel voor het schaambeen. Het dijbeen heeft een bult op de binnenste beenstijl naar de binnenste onderste gewrichtsknobbel. De groeve tussen de onderste gewrichtsknobbels van het dijbeen is naar onderen open. De bult op de beenstijl naar de binnenste onderste gewrichtsknobbel is naar achteren gericht. Het vierde middenvoetsbeen heeft een buitenste richel voor de hechting van de buigende spieren.

Bij het bovenstaande moet bedacht worden dat de beschrijvers aanhangers zijn van de hypothese van de frame shift bij de telling van de vingers. Ze houden zich dus aan de traditionele identificatie, op basis van embryologie, van de drie vingers van huidige vogels als de tweede, derde en vierde vinger van de Amniota. Ze erkennen echter dat, gezien de gevonden fossielen, de vroegste theropode voorouders van de vogels op deze posities de eerste, tweede en derde vinger hadden. Een genetische verschuiving zou de structuur van de eerste, tweede en derde vinger overgebracht hebben naar de tweede, derde en vierde vinger. Dit zou dan gebeurd zijn aan de basis van de Maniraptora waartoe ook Manipulonyx behoort. Veel westerse paleontologen menen echter dat er helemaal geen frame shift geweest is en dat ook moderne vogels de eerste, tweede en derde vinger in de handen hebben. Wat hierboven dus als eerste vinger of duim wordt aangeduid, noemen de beschrijvers de tweede vinger, en de kleine vingers zijn dan niet de tweede en derde, maar de derde en vierde.
Skelet
Het meest opvallende deel van het skelet zijn de voorpoten die vollediger zijn dan ooit eerder bij de Alvarezsauridae aangetroffen. Juist vanwege die grotere compleetheid is een vergelijking met verwanten lastig omdat niet kan worden vastgesteld of die een bepaald kenmerk ook bezitten. Niet eerder waargenomen eigenschappen kunnen dus uniek zijn voor Manipulonyx of juist kenmerkend voor de groep als geheel.
De enorme duimklauw wordt met veel verwanten gedeeld. De veel kleinere tweede en derde vinger zijn echter niet gedegenereerd in structuur maar hebben ieder drie kootjes waaronder een eindklauw. De middenhandsbeenderen zijn versmolten tot één enkele enorme structuur, breder dan de hele pols en de uiterste polsbeenderen met holle kassen omklemmend. De tweede en derde vinger steken daar zijwaarts uit. De eerste vinger steekt naar voren, met de basis verbreed tot driekwart van de breedte van de middenhand, rustend op twee voorste knobbels daarvan.
Op de palmzijde van de hand bevinden zich drie huidverbeningen of osteodermen, een situatie die van directe verwanten niet bekend is. Überhaupt zijn onder de Theropoda verder alleen bij Ceratosaurus osteodermen aangetroffen maar dan op de rug. Die op de hand van Manipulonyx hebben dezelfde vorm als bij de Sauropoda en de Ornithischia, met een centrale kiel waarop een hoogste punt. De osteodermen hebben een verschillende grootte. De kleinste ligt tussen de eerste vinger en de tweede. Van de twee grotere osteodermen ligt er een op de binnenzijde van het middenhandsbeen, als een duimpje uitstekend. De andere ligt midden op de handpalm.
Fylogenie
Manipulonyx is binnen de Alvarezsauridae in de Parvicursorinae geplaatst maar zonder exacte kladistische analyse.
Levenswijze
Van Alvarezsauridae is vaak gedacht dat ze zich met hun grote duimklauw en spitse snuit voedden met mieren of termieten in een gravende levenswijze. Deze hypothese werd vooral begin jaren negentig voorgestaan door John Ostrom. De beschrijvers van Manipulonyx wezen op kenmerken die daarmee niet in overeenstemming waren. De binnenste beenstijl van het opperarmbeen is zwak ontwikkeld. Om te graven is het handig om de handpalm naar de grond te richten maar tot een dergelijke pronatie was de hand niet in staat. De hand is ook niet gevormd als een spade, "skalodactiel", als bij gravende zoogdieren. Verder werd gewezen op de wanverhouding in lengte tussen de armen en achterpoten. Nu zijn de lange achterpoten nog niet zo bezwaarlijk voor een myrmycofage vorm. Die hoeft geen heel hol te graven en dus niet losgewerkte aarde niet met de voeten weg te werken. Daarbij zou het voor de kleine alvarezsauriden wezenlijk zijn geweest zich op de lange acherpoten met een snelheid van ruim 50 km/h uit de voeten te hebben kunnen maken voor de grote theropoden in hun leefgebied. Een probleem zouden echter de korte armen geweest zijn want die konden nauwelijks de grond raken zonder op de buik te gaan liggen. Daarom is wel aangenomen dat ze in hoge termietennesten probeerden te breken of de bast van bomen poogden te trekken om keverlarven te bereiken.
Eind twintigste eeuw viel steeds meer onderzoekers op dat vaak eierschalen van kennelijk andere theropoden samen met alvarezsauride fossielen werden aangetroffen. Dat suggereerde dat die eierrovers waren, wat ook al eens gedacht was van Oviraptor (die daarom "eierrover" gedoopt was), totdat bleek dat die op haar eigen eieren zat. Toch was dit een plausibel idee want de wijfjes van grote dinosauriërs produceerden enorme legsels van tientallen eieren, ieder soms met ruim een liter inhoud, een aanzienlijk potentiële energiebron. Het probleem is dat die gelijktijdig gelegd werden tijdens een relatief kort broedseizoen en alvarezsauriden te klein waren om een grote vetreserve aan te leggen, wat de vraag oproept waarvan zij dan de rest van het jaar leefden. Desalniettemin wisten de beschrijvers een aantal kenmerken aan te geven die wezen op het roven van eieren.
De Parvacursorinae hebben zijwaarts gerichte schoudergewrichten. Daardoor konden de opperarmbeenbeenderen goed naar bezijden uitsteken om een groot ei te omklemmen. De Parvacursorinae hebben een sterk ontwikkelde deltopectorale kam op het opperarmbeen en een krachtige kiel op de gepaarde borstbeenderen. Tussen beide structuren kan een krachtige musculus pectoralis major gelopen hebben, een spier die de arm naar voren en binnen trok, geschikt om een ei vast te houden. Het opperarmbeen had één in plaats van twee gewrichtsknobbels met de onderarm, een klein bol ovaal vlak vormend aan de achterzijde. Dit paste in een ovale uitsparing in de vergroeide ellepijp en spaakbeen. Dit beperkt de beweging tot een enkelvoudige strekking en buiging. Er is een grote processus olecrani, een hefboom om de arm in een plotse ruk via de musculus triceps bracchii te strekken, wat gediend kan hebben om de hand los te rukken uit een ei om dat op de grond open te laten barsten teneinde de inhoud op te slurpen. Een vierde kenmerk is de vorm van de hand met stekels en kleine zijvingers. De stekels zijn alleen in Manipulonyx bekend maar de vorm van de middenhand bij verwanten duidt er volgens de beschrijvers op dat althans de stekel op de binnenzijde aanwezig was. De hoornschacht op de stekels zou gebruikt kunnen zijn om glibberige eieren vast te houden en ze te kraken. Een vijfde aanwijzing is de sterk ontwikkelde buitenstijl van het opperarmbeen, de aanhechting voor een krachtige musculus anconeus, die ook weer de onderarm strekt. Een laatste bewijs is het ontbreken op de duimklauw van een bult voor de aanhechting van de pees van de buigende spier. Dat past niet bij een gravende functie omdat het daarbij juist belangrijk is de klauwen te krommen. De bult dreigt echter vast te komen zitten in een eierschaal en dat zou het verdwijnen ervan kunnen verklaren.
Al deze zaken overwegende hebben de beschrijvers een scenario opgesteld over de levenswijze. Uit de ogen die dieptezicht zouden hebben toegestaan en de scherpe oren leidden ze af dat het een nachtdier betreft. In de nacht zouden de dieren nesten beslopen hebben om eieren te roven. Een enkel groot ei omklemmend zouden ze zich met de lange achterpoten uit de voeten hebben gemaakt. Dat zou er op wijzen dat de dinosauriërs broedzorg kenden en de dieven probeerden te achterhalen. Soms zou dat ook gelukt zijn en dat zou de combinatie van alvarezsauride skeletten met eierschalen verklaren. De hypothese van Qin dat de dieren zo klein waren doordat ze mieren aten verwerpen de beschrijvers want mierenetende zoogdieren namen ook niet in omvang af. De geringe omvang zou een aanpassing zijn aan de grootte van de eieren.
Literatuur
- A.O. Averianov, A.V. Lopatin and A.A. Atuchin. 2025. "Forelimb structure and function in a new Late Cretaceous parvicursorine theropod dinosaur from Mongolia". Proceedings of the Zoological Institute RAS. 329(4): 382–408. DOI:10.31610/trudyzin/2025.329.4.382.