Dinocephalosaurus
| Dinocephalosaurus Status: Uitgestorven, als fossiel bekend Fossiel voorkomen: Midden-Trias | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||
| Taxonomische indeling | ||||||||||||||
| ||||||||||||||
| Geslacht | ||||||||||||||
| Dinocephalosaurus Li, 2003 | ||||||||||||||
| Typesoort | ||||||||||||||
| Dinocephalosaurus orientalis | ||||||||||||||
| Afbeeldingen op | ||||||||||||||
| Dinocephalosaurus op | ||||||||||||||
| ||||||||||||||
Dinocephalosaurus[1][2][3][4] ('reptiel met een vreeswekkende kop') is een geslacht van uitgestorven langhalzige aquatische Prolacertiformes met een uitzonderlijk lange nek met vijfentwintig wervels, dat de zeeën uit het Midden-Trias van het huidige China bewoonde, behorend tot de Archosauromorpha. De leeftijd van de fossielen is vastgesteld op ongeveer 245 miljoen jaar. Vertegenwoordigers van dit geslacht zwommen in het Midden-Trias door de zeeën van het huidige Zuid-China. Het geslacht bevat het type en de enige bekende soort Dinocephalosaurus orientalis, die in 2003 door Li werd benoemd. In tegenstelling tot andere Prolacertiformes met lange nek (die een groep vormen die bekend staat als de Tanystropheidae), ontwikkelde Dinocephalosaurus convergent een lange nek, niet door verlenging van individuele halswervels, maar door toevoeging van halswervels die op zich een gemiddelde lengte hebben. Net als andere tanystropheïden gebruikte Dinocephalosaurus waarschijnlijk zijn lange nek om op prooien te jagen, met behulp van een combinatie van zuigkracht, gecreëerd door de uitzetting van de keel, en lange vangtanden van de kaken om prooien te grijpen. Het was waarschijnlijk een puur zeedier, zoals gesuggereerd door de slecht verbeende en peddelachtige ledematen die zouden hebben voorkomen dat het aan land ging.
Exemplaren die tot het geslacht behoren, werden voor het eerst ontdekt in 2002 in een plaats nabij Xinmin in Guizhou. Op dezelfde vindplaats, die dateert van 244 miljoen jaar geleden, zijn ook andere mariene reptielen gevonden, zoals Mixosaurus, Keichousaurus en Wumengosaurus. Terwijl het type-exemplaar alleen uit een schedel en de voorkant van de nek bestond, onthulden andere exemplaren al snel de volledige vorm van het lichaam. Verdere ontdekkingen van Dinocephalosaurus-exemplaren werden vanaf 2008 gedaan in Luoping, Yunnan. Op deze locatie zou Dinocephalosaurus naast Mixosaurus, Dianopachysaurus en Sinosaurosphargis hebben geleefd. Een exemplaar dat op de plaats Luoping werd ontdekt, had een foetus in zijn buik, wat aangeeft dat Dinocephalosaurus levendbarend was zoals veel andere mariene reptielen. Dinocephalosaurus is het enige bekende lid van de Archosauromorpha dat levendbarend is, met mogelijke uitzondering van de Metriorhynchidae, een groep mariene Crocodylomorpha.
Ontdekking en naamgeving
Panxian
.png)
Het type-exemplaar of holotype van Dinocephalosaurus werd ontdekt in 2002 tijdens veldwerk, uitgevoerd bij het dorp Yangjuan, Xinmin District, in de prefectuur Panxian, in Guizhou. Het bestaat uit een bijna volledige schedel die de linkerkant van de kaak mist, evenals verschillende bijpassende halswervels. Het werd vervolgens opgeslagen in het Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) te Beijing onder het inventarisnummer IVPP V13767. Het beschrijvende artikel werd door IVPP-paleontoloog Li Chun gepubliceerd in Acta Geologica Sinica in december 2003.[5]
Een tweede specimen dat op dezelfde vindplaats werd ontdekt, vertegenwoordigt een gedeeltelijk gearticuleerd skelet dat alleen de staart mist. Eveneens opgeslagen bij het IVPP heeft het exemplaar het inventarisnummer IVPP V13898. Het exemplaar werd toegewezen en beschreven in een kort artikel door Li, Olivier Rieppel[6] en Michael LaBarbera, dat in september 2004 in Science werd gepubliceerd.[7] Een meer gedetailleerde beschrijving werd vervolgens gepubliceerd door Rieppel, Li en Nicholas Fraser[8] in een Journal of Vertebrate Paleontology artikel uit 2008.[9]
Toegewezen zijn verder de specimina IVPP V17977: een kop en een nek; IVPP V20295: een volledig skelet; ZMNH M8727: de voorste helft van een skelet; ZMNH M8728: een voorste skelet; en ZMNH M8752: een volledig maar sterk samengedrukt skelet.
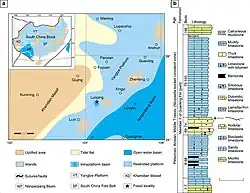
De vindplaats Panxian, waar deze exemplaren vandaan kwamen, maakt deel uit van Afzetting II van de Guanlingformatie uit het Anisien (Midden-Trias), waarvan aanvankelijk werd gedacht dat deze 230 miljoen jaar oud was, maar het meest recent is gedateerd op 244 ± 1,3 miljoen jaar oud op basis van uranium-looddatering. De overheersende afzettingen op deze plaats zijn samengesteld uit grijze tot donkergrijze mergelkalksteen, evenals cherty-kalksteen met dolomiet- en bentonietbedden. Dinocephalosaurus werd specifiek gevonden in laag 90 van de Panxianafzettingen, een dunne kalksteenlaag die traditioneel wordt toegewezen aan de bovenste Reptile Horizon (lagen 87-90). Lager liggen de Middle Reptile (lagen 81-85) en Lower Reptile (lagen 77-79) Horizons.
Luoping
Vervolgens werden aanvullende Dinocephalosaurus-exemplaren ontdekt uit de iets oudere vindplaats Luoping, die is gedateerd op 245-244 miljoen jaar oud op basis van conodont-biostratigrafie en voorlopige radiometrische datering. De specimina zijn afkomstig uit bed 74 van de vindplaats Luoping, in afzettingen in de buurt van het dorp Dawazi, Luoping County, Yunnan. Bed 74 maakt deel uit van een sectie, bestaande uit dunne donkergrijze micritische kalksteenlagen, vermengd met dikkere lagen kiezelhoudende kalksteen en siltige kalksteen, die zich naar beneden uitstrekt tot bed 67.
Een speciaal exemplaar is opmerkelijk omdat het een embryo in zijn buikstreek bevat, waarvan halswervels, voorpoten en verschillende andere elementen bewaard zijn gebleven. Het werd verzameld in 2008. Toen had verwering het exemplaar al in drie blokken gesplitst, waarbij de spleten waren opgevuld door hedendaagse aarde. Het exemplaar werd vervolgens overgebracht naar het Chengdu Center van de China Geological Survey, waar het werd geprepareerd en opgeslagen onder het inventarisnummer LPV 30280. Vervolgens werd het exemplaar beschreven door Liu Jun, Chris Organ, Michael Benton, Matthew Brandley, en Jonathan Aitchison, in februari 2017 in Nature Communications.
Een verder specimen werd in 2013 gemeld, ZMNH M8752.[10]
Naamgeving
In 2003 benoemde Li de typesoort Dinocephalosaurus orientalis. De geslachtsnaam is een combinatie van deinos, "vreeswekkend", kephalè, "hoofd", en sauros, "hagedis". Volgens Li verwijst deze naam naar de 'afschuwelijke schedel' van het holotype. De soortaanduiding is het Latijn orientalis, "oostelijk", een verwijzing naar de status als enige bekende soort van de Tanystropheidae uit het oostelijke deel van de Tethysoceaan.
Kenmerken

Dinocephalosaurus was een groot lid van de Protorosauria en bereikte een lichaamslengte van minstens 3,5 meter, vergeleken met een maximum van drie meter voor Tanystropheus. De bekende exemplaren zijn waarschijnlijk volwassen, aangezien ze versmolten schedelbotten hebben en de spits toelopende tanden missen, die te zien zijn bij juveniele leden van Tanystropheus. Net als Tanystropheus heeft Dinocephalosaurus een uitzonderlijk lange nek (1,7 meter lang) ten opzichte van zijn romp (een meter zonder de staart).
Schedel
De schedel van Dinocephalosaurus is laag en smal, met een lange premaxilla en maxilla vergeleken met die van Tanystropheus. Zowel de premaxilla als de maxilla ontmoeten elkaar in de voorste onderste hoek en dragen bij aan de rand van het neusgat, dat zich aan het voorste uiteinde van een lange uitsparing bevindt, die zich langs de snuit voor de oogkas uitstrekt (de antorbitale depressie). De onderranden van de twee bovenkaakbotten per zijde dragen respectievelijk vijf en twaalf lange en spitse tanden; de derde premaxillaire en vierde en vijfde maxillaire tanden zijn duidelijk verlengd. De onderkaak heeft vijftien bewaarde tanden, waarvan er drie slagtandachtig zijn. Ter vergelijking: de tanden bij volwassen exemplaren van Tanystropheus zijn scherpe kegels, terwijl ze tricuspide zijn (met drie knobbels) bij jonge exemplaren.

In tegenstelling tot de ovale oogkas van Tanystropheus, lijkt de oogkas van Dinocephalosaurus perzikvormig, met een smal voorste uiteinde. Op de bovenkant van de schedel is het wandbeen breed en afgeplat, zonder sporen van de middenlijnkam die Tanystropheus toont. Het jukbeen heeft slechts twee takken, waarbij de derde achterste tak ontbreekt die aanwezig is bij de meeste andere reptielen. Ook ontbreekt het retro-articulaire uitsteeksel van de achterste onderkaak (een ander onderscheidingspunt met Tanystropheus), evenals extra tanden op en een holte tussen de pterygoide botten van het verhemelte.
Nek en romp
.png)
Tanystropheus en Dinocephalosaurus bereikten hun extreem langgerekte nek op verschillende manieren. De nek van Tanystropheus bestaat uit dertien langwerpige halswervels, terwijl de hals van Dinocephalosaurus bestaat uit ten minste zevenentwintig halswervels die op zich niet zo langwerpig zijn. Van de zevenentwintig wervels van Dinocephalosaurus is de negentiende de langste, die ongeveer eenennegentig millimeter lang is. Ter vergelijking: de langste wervels in een Chinees exemplaar van Tanystropheus zijn de negende en tiende, die 248 millimeter lang zijn. Bovendien zijn de halswervels van Dinocephalosaurus niet hol, in tegenstelling tot die van Tanystropheus.
.png)
Bijkomende kenmerken van de halswervels die Dinocephalosaurus onderscheiden van andere protorosauriërs zijn de lage en kielachtige doornuitsteeksels en de voorste en achterste gewrichtsoppervlakken van de wervels, die beide hol gekromd concaaf zijn. Bij de eerste tien wervels is de onderrand ook hol. De lange, slanke halsribben dragen voorste uitsteeksels die vrij zijn van de wervellichamen, en die ook uniek zijn voor Dinocephalosaurus. Deze ribben zijn uitgelijnd langs de nek en overbruggen meerdere opeenvolgende wervelgewrichten, van twee of drie opeenvolgende gewrichten in de voorste nek tot vijf of zes in de achterste nek. Er lijken geen duidelijke uitsteeksels op de wervels te zijn voor articulatie met de ribben.
De romp van Dinocephalosaurus heeft minstens zesentwintig wervels. Bij Dinocephalosaurus lijken er geen lendenwervels te zijn in de zin van wervels van de romp zonder ribben. De ribben van het heiligbeen en de staart lijken ook niet te zijn versmolten met hun corresponderende wervels. Elk van de gastralia (buikribben) bij Dinocephalosaurus bestaat uit drie overdwars geplaatste elementen in plaats van vier zoals bij Tanystropheus. Ze verschillen doordat Dinocephalosaurus slechts één centraal element op de middenlijn heeft, terwijl Tanystropheus twee elementen heeft die samen een middenlijnstaaf vormen.
Ledematen
Dinocephalosaurus had relatief grote poten, die eindigden in flipperachtige handen en voeten. De voor- en achterpoten zijn ongeveer even lang, in tegenstelling tot Tanystropheus, waar de voorpoten veel kleiner zijn. Terwijl de meeste protorosauriërs, zoals Tanystropheus, Macrocnemus en Langobardisaurus, relatief verbeende ledematen hadden die waren aangepast voor het leven op het land, zijn de stevige ledematen van Dinocephalosaurus slecht verbeend en lijken ze op die van nothosauriërs. Van de carpale botten in de pols zijn er slechts zes verbeend, overeenkomstig zijn slechts drie van de tarsale botten in de enkel verbeend. Bovendien articuleren het sprongbeen en het hielbeen ook niet met elkaar in de enkel, in plaats daarvan vormen ze eenvoudige en ronde ossificaties, verbeningen.
Deze eigenschappen zijn waarschijnlijk neoteen, zoals bij veel andere aquatische tetrapoden. Tanystropheus is waarschijnlijk ook neoteen is, maar niet in de mate van Dinocephalosaurus. Verschillende andere eigenaardige eigenschappen zijn aanwezig in de voeten van Dinocephalosaurus. In tegenstelling tot Tanystropheus en de meeste andere protorosauriërs, is het vijfde middenvoetsbeentje van Dinocephalosaurus eenvoudig en recht in plaats van haakvormig. Tanystropheus heeft slechts een vijfde middenvoetsbeentje dat aan de bovenkant licht verdikt is en het heeft bovendien een lang kootje op de vijfde teen dat niet anders is dan bij de andere tenen. Op de derde teen van de voet zijn er vier kootjes, maar geen daarvan lijkt de eindklauw te zijn, wat suggereert dat Dinocephalosaurus minstens vijf kootjes in de derde teen had, een hoger dan gemiddeld aantal.
Fylogenie
Dinocephalosaurus wordt beschouwd als een lid van de Protorosauria, een groep wijd verspreide en diverse reptielen uit het Perm-Trias. Deze toewijzing is gebaseerd op kenmerken als de lage en smalle schedel met een korte en smalle postorbitale regio, het lange neusbeen ten opzichte van het voorhoofdsbeen, de gereduceerde achterste tak van het jukbeen, de aanwezigheid van meer dan zeven halswervels, met centra langer dan die van de ruggenwervels, de lage doornuitsteeksels van de halswervels, de lange nekribben, het ontbreken van intercentra die articuleren met de dorsale centra en de afwezigheid van een foramen entepicondylare op het opperarmbeen. Deze kenmerken worden gedeeld met Tanystropheus, Macrocnemus en andere protorosauriërs.
Protorosauriërs werden vroeger beschouwd als basale Lepidosauromorpha, maar fylogenetische analyse heeft vervolgens aangetoond dat het in feite niet-archosaurische archosauromorfen zijn. Zoals oorspronkelijk gedefinieerd, verwees de Protorosauria naar de groep met Protorosaurus en Prolacerta en was synoniem met de Prolacertiformes. In 1997 voerde Nour-Eddine Jalil een analyse uit van de Prolacertiformes, die de definitie uitbreidde tot veertien geslachten, waaronder de Tanystropheidae. Daaropvolgend onderzoek heeft over het algemeen aangetoond dat Prolacerta dichter bij de Archosauriformes staat dan Protorosaurus, waardoor de Prolacertiformes in de traditionele zin niet-monofyletisch zijn. Of de resterende protorosauriërs, inclusief Protorosaurus, tanystropheiden en drepanosauriërs een monofyletische groep vormen, is onduidelijk. Verschillende fylogenieën hebben zowel monofyletische als niet-monofyletische Protorosauria ondersteund, in het laatste geval met Protorosaurus als meer basaal dan de Tanystropheidae.
In de oorspronkelijke beschrijving van Dinocephalosaurus herkende Li de zone achter de oogkas en de langwerpige halswervels als een aanwijzing voor een nauwe verwantschap met Tanystropheus; hij plaatste het dier in de Tanystropheidae. Rieppel et alii merkten vervolgens echter kenmerken op die Dinocephalosaurus onderscheidden van afgeleide protorosauriërs als Tanystropheus, Macrocnemus, Langobardisaurus en Tanytrachelos. Deze omvatten het ontbreken van een taps toelopende voorkant van het neusbeen en een niet-gereduceerde bijdrage van het darmbeen aan de heupkom. Dit bracht Rieppel et alii ertoe om Dinocephalosaurus te beschouwen als een onbepaalde protorosauriër, met zijn nekverlenging convergerend met die van Tanystropheus. Hun fylogenetische stamboom, gebaseerd op een dataset afgeleid van de afzonderlijke analyses van Jalil (1997), David Dilkes (1998) en Michael Benton & Jackie Allen (1997), wordt hieronder weergegeven.
Topologie A: Rieppel et alii (2008)
| Protorosauria |
| ||||||||||||||||||||||||
Topologie B: Liu et alii (2017)
| Protorosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Liu et alii voerden in 2017 een afzonderlijke fylogenetische analyse uit. Ze bekritiseerden de analyse van Rieppel et alii omdat ze meerdere kenmerken in hun gegevensverzameling meermalen hadden opgenomen, in iets andere omschrijvingen die elkaar impliceren, waardoor de dubbel getelde kenmerken een te groot gewicht kregen in de analyse. In hun eigen analyse gebruikten Liu et alii dezelfde brondatasets, maar verwijderden dubbel getelde kenmerken, voegden twee nieuwe kenmerken toe uit een analyse door Sean Modesto en Hans-Dieter Sues (2004) en verwijderden slecht bewaarde of mogelijk chimaere taxa zoals zoals Cosesaurus, Kadimakara, Trachelosaurus en Malerisaurus. Analyses op basis van op spaarzaamheid gebaseerde en bayesiaanse methoden vonden dat Dinocephalosaurus een lid was van de Tanystropheidae, de zustergroep van een clade met Tanystropheus, Macrocnemus, Langobardisaurus en Tanytrachelos. De analyses leverden ook een monofyletische Protorosauria op, zij het met uitsluiting van Prolacerta. De stamboom uit de op spaarzaamheid gebaseerde analyse is hierboven weergegeven, rechts.
Paleobiologie
Nek en voeding
De lange nek van Dinocephalosaurus heeft waarschijnlijk een functionele rol gespeeld. Hij zorgde voor een grote afstand tussen het hoofd en de rest van het lichaam. Dit zou Dinocephalosaurus in staat hebben gesteld potentiële prooien te naderen zonder dat het grootste deel van zijn romp werd waargenomen, wat nuttig zou zijn geweest in de troebele wateren van zijn leefgebied.
Dinocephalosaurus heeft mogelijk ook zijn nek gebruikt om zijn prooi te vangen via zuigvoeding. Na het opzij buigen van de nek (wat vergemakkelijkt zou zijn door de slankheid van de nekribben), zou het rechttrekken van de nek ertoe hebben geleid dat de nekribben naar buiten werden gespreid door de werking van de nekspieren die aan de ribben waren bevestigd. Naarmate het hoofd naar voren sprong, zou het volume van de slokdarm zijn toegenomen, waardoor zuigkracht ontstond. Zodra de prooi was gevangen, zouden de hoektanden de prooi in de bek hebben vastgehouden. Net als andere aquatische amnioten, zou de prooi van Dinocephalosaurus met de kop naar voren zijn ingeslikt en verteerd, zoals blijkt uit de wijze waarop een perleidiforme vis in de buikholte van LPV 30280 uit Luoping bewaard is.
Het is niet waarschijnlijk dat Dinocephalosaurus zijn lange nek gebruikte om te ademen door hem verticaal uit te strekken. Als het dat zou proberen, zou het drukverschil tussen het oppervlak en zijn romp te groot zijn om zijn longen te kunnen vullen. Dus om te ademen, zou Dinocephalosaurus het oppervlak moeten naderen met een bijna horizontale nek. De horizontale houding van de nek zou overigens de voortbeweging aan het oppervlak niet hebben vergemakkelijkt, omdat het lange profiel dat de romplengte vergroot de waterweerstand juist zou hebben verhoogd.
Voortplanting

Dinocephalosaurus vertegenwoordigt het hoogtepunt van aquatische aanpassingen onder de protorosauriërs. Gezien zijn lange nek en peddelachtige ledematen was hij waarschijnlijk niet in staat om zonder ongemak op het land te lopen. Een gevolg van deze aanpassingen is dat Dinocephalosaurus niet in staat zou zijn geweest om nesten op het land te bouwen. Dit zou ook hebben verhinderd dat hij reptieleneieren met een harde of zelfs leerachtige schaal legde. Dergelijke eieren vereisen de uitwisseling van gassen met de buitenomgeving en dit proces is aanzienlijk langzamer in water dan in lucht. Dinocephalosaurus kan dus niet ovipaar ofwel eierleggend zijn geweest. Tegelijkertijd staan archosauriërs echter bekend om de totale afwezigheid van levendbarende moderne leden, namelijk de vogels en de krokodilachtigen.
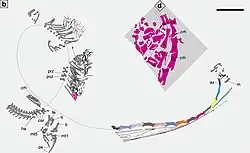
Het embryonale individu van Dinocephalosaurus, bewaard in specimen LPV 30280 uit Luoping, kan om verschillende redenen als zodanig worden geïdentificeerd. Ten eerste is het volledig ingesloten in de lichaamsholte van de volwassene. De lange nekribben, als die van de volwassene, wijzen in dezelfde richting als de ruggenwervels van de volwassene, wat in tegenspraak is met de typische methode om prooien onder amnioten in te slikken. Het is ook bewaard met een nek die naar zijn voorpoten krult, een houding die typisch wordt gezien bij embryo's van gewervelde dieren, maar niet bij volwassen exemplaren van Dinocephalosaurus, die over het algemeen de nek naar boven hebben gebogen. De afwezigheid van handen in anatomisch verband met de voorpoten in het embryo kan een artefact van de bewaring zijn, maar het kan ook te wijten zijn aan de volgorde waarin de ledematen van tetrapoden verbenen, namelijk met de uiteinden het laatst.
Hoewel de aanwezigheid van een embryo hierdoor nog niet dwingend bewezen wordt, leveren de omstandigheden waarin het embryo werd bewaard sterk bewijs dat Dinocephalosaurus levendbarend was. Er is geen bewaarde verkalkte eierschaal rond het embryo, ondanks de aanwezigheid van fijn bewaarde verkalkte elementen van andere dieren in Luoping. Dit komt overeen met het gegeven dat de eierschalen die de embryo's van levendbarende reptielen omringen, niet-verkalkte membranen zijn. Bovendien geven de relatieve verhoudingen van het opperarmbeen en het kuitbeen in het embryo, vergeleken met het moederdier en specimen IVPP V13898 van Panxian, aan dat het embryo ongeveer twaalf procent van de lengte van zijn moeder heeft. In combinatie met zijn verbeende botten suggereert dit dat het embryo zich in een vergevorderd ontwikkelingsstadium bevond, terwijl krokodilachtigen, vogels, schildpadden en tuatara's eieren leggen met embryo's in zeer vroege ontwikkelingsstadia.
Over het algemeen suggereert het door het embryo geleverde bewijs dat Dinocephalosaurus levendbarend was, waardoor het de eerste levendbarende archosauromorf zou zijn, afgezien van mogelijk metriorhynchiden. Dit komt overeen met de scheiding van zijn sacrale ribben van zijn heiligbeen, wat wijst op een beweeglijk bekken, wat de bevalling zou hebben kunnen vergemakkelijkt. Hoewel de verdere afwezigheid van levendbarende archosauriërs in het verleden is toegeschreven aan gemeenschappelijke kenmerken die zijn geërfd van de archosaurische stamlijn, suggereert de ontdekking dat Dinocephalosaurus levendbarend was dat dit fenomeen niet te wijten is aan afstammings-specifieke levensstijlbeperkingen. Hoewel de geslachtsbepalingssystemen onder levende archosauromorfen divers zijn, met krokodilachtigen en schildpadden die temperatuurafhankelijke geslachtsbepaling gebruiken, suggereert fylogenetische modellering dat Dinocephalosaurus de basale toestand van genotypische geslachtsbepaling door geslachtschromosomen van vroege diapsiden behield en dat dit systeem de overgang ervan vergemakkelijkte tot een verplichte mariene levensstijl gekoppeld aan levendbarendheid.
Paleo-ecologie
Tot tenminste het einde van het Midden-Trias zorgde de hoge zeespiegel ervoor dat ondiep water een groot deel van het Zuid-Chinese blok bedekte, een tektonische plaat die tegenwoordig bestaat uit het stabiele Yangtze-Craton en de minder stabiele Zuid-Chinese verschuiving. Een gebergtevormingsgebeurtenis, bekend als de Indochinese orogenese, heeft Precambrische rotsen opgetild om vier grote landmassa's op het Zuid-Chinese blok te vormen: Khamdian in het westen, Jiangnan in het centrum, Yunkai in het zuiden en Cathaysia in het oosten. Eilandketens strekten zich ook uit tussen Yunkai en Cathaysia in het oosten. Gelegen tussen Khamdian en Jiangnan was een diep oceaanbekken bekend als het Nanpanjiang Basin. Langs de westelijke rand van dit bassin werden fossielhoudende sedimenten afgezet om te worden wat nu de Lagerstätten zijn (sedimentaire afzettingen die worden gekenmerkt door een voorbeeldige bewaring) van Panxian, Luoping en andere vindplaatsen.
Bij Panxian lijkt er een overgang te zijn geweest in de endemische mariene reptielenfauna, mogelijk veroorzaakt door vulkanisme. Dit vulkanisme wordt aangegeven door de aanwezigheid van een bentonietlaag tussen de middelste en hogere Reptile Horizon. In de lagere en de bovenste Reptile Horizon zijn ook fossielen gevonden van de alomtegenwoordige mixosauride Mixosaurus panxianensis (die in alle lagen voorkomt), de pachypleurosauriden Keichousaurus sp. en Wumengosaurus delicatomandibularis en vissen, hoewel fossielen van de laatste fragmentarisch zijn. De lagere middelste- en lagere Reptile Horizon omvatten ook de mixosauride Phalarodon cf. fraasi, de basale ichthyosauriër Xinminosaurus catactes, de placodont Placodus inexpectatus, de nothosauriërs Lariosaurus hongguoensis en Nothosaurus yangjuanensis, en de archosauriër Qianosuchus mixtus, naast tweekleppigen en saurichthyide vissen.
De fauna van Luoping lijkt te zijn bewaard in een klein bassin binnen het platform in plaats van in het omringende open water, te oordelen naar de anoxische sedimenten die op de locatie aanwezig zijn. Van de 19 759 exemplaren bestaat 93,7 procent van de fossielen die in Luoping zijn gevonden uit geleedpotigen: tienpotigen, isopoden, krabachtige Cyclida, garnaalachtige mysidaceeën, mosselgarnalen en ostracoden, evenals zeldzame duizendpoten en hoefijzerkrabben. Ter vergelijking: slechts 0,07 procent van de exemplaren is afkomstig van mariene reptielen, waaronder Dinocephalosaurus naast Mixosaurus cf. panxianensis, Dianopachysaurus dingi, Sinosaurosphargis yunguiensis en een archosauriër. Verder zijn er vissen, waaronder saurichthyiden, paleonisciden, birgeriiden, perleididen, eugnathiden, semionotiden, pholidopleuriden, peltopleuriden en coelacanthiden gevonden in Luoping, die 3,66 procent van de fossielen vormen met vijfentwintig taxa in negen families. Weekdieren zoals tweekleppigen en gastropoden, samen met ammonoïden en belemnoïden, zijn goed voor 1,69 procent van de exemplaren. Zeldzame en waarschijnlijk niet-endemische fossielen omvatten ook stekelhuidigen zoals crinoïden, zeesterren en zee-egels, branchiopoden en relatief complete naaldboomtakken en bladeren, die waarschijnlijk afkomstig zijn uit kustbossen op minder dan tien kilometer afstand.
- ↑ Dinocephalosaurus. www.prehistoric-wildlife.com. Gearchiveerd op 13 november 2021. Geraadpleegd op 13-11-2021.
- ↑ Mindat.org. www.mindat.org. Geraadpleegd op 13-11-2021.
- ↑ Fossilworks: Dinocephalosaurus. fossilworks.org. Gearchiveerd op 13-11-2021. Geraadpleegd op 13-11-2021.
- ↑ (en) Mike, First Live Birth Evidence in Ancient Dinosaur Relative. Everything Dinosaur Blog. Geraadpleegd op 13-11-2021.
- ↑ Li, C. 2003. "First record of protorosaurid reptile (Order Protorosauria) from the Middle Triassic of China". Acta Geologica Sinica - English Edition. 77(4): 419–423
- ↑ (en) Staff Profile. Field Museum. Gearchiveerd op 13-11-2021. Geraadpleegd op 13-11-2021.
- ↑ Li, C.; Rieppel, O. & LaBarbera, M.C. 2004. "A Triassic Aquatic Protorosaur with an Extremely Long Neck". Science. 305(5692): 1931
- ↑ (en) Dr Nick Fraser. National Museums Scotland. Geraadpleegd op 13-11-2021.
- ↑ Rieppel, O.; Li, C. & Fraser, N.C. 2008. "The Skeletal Anatomy of the Triassic Protorosaur Dinocephalosaurus orientalis Li, from the Middle Triassic of Guizhou Province, Southern China". Journal of Vertebrate Paleontology. 28(1): 95–110
- ↑ Fraser, N.C.; Rieppel, O. & Li Chun. 2013. "A long-snouted protorosaur from the Middle Triassic of southern China". Journal of Vertebrate Paleontology. 33(5): 1120–1126
- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Dinocephalosaurus op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.